Anda mungkin juga menyukai
- Método de Escultura em Bloco de Cera-Incisivo Central Su Perior-21Dokumen15 halamanMétodo de Escultura em Bloco de Cera-Incisivo Central Su Perior-21Cleisson Sampaio91% (11)
- Plano de treinamento PRFDokumen7 halamanPlano de treinamento PRFlcnferreiraBelum ada peringkat
- Resumo de Imunologia abrangendo os principais conceitosDokumen29 halamanResumo de Imunologia abrangendo os principais conceitosÍtalo AlmeidaBelum ada peringkat
- Exercícios 1a Lei MendelDokumen6 halamanExercícios 1a Lei MendelRafael Fernandes100% (1)
- Sistema imune: respostas inata e adaptativaDokumen8 halamanSistema imune: respostas inata e adaptativamarcial_pinazoBelum ada peringkat
- Citocinas - ImunologiaDokumen4 halamanCitocinas - ImunologiaAluno - Polyana Aparecida Bisinoto100% (1)
- Hipersensibilidade Tipo 3 (Trabalho Final)Dokumen3 halamanHipersensibilidade Tipo 3 (Trabalho Final)saulopaulino100% (1)
- Introdução A ImunologiaDokumen31 halamanIntrodução A ImunologiaSol MontosaBelum ada peringkat
- Resumo Sobre Documentário: O Poder Do CérebroDokumen3 halamanResumo Sobre Documentário: O Poder Do CérebroHellayne Sousa100% (1)
- Apostila Dry Needling Golden CursosDokumen44 halamanApostila Dry Needling Golden CursosAMANDA CLARA100% (1)
- Apostila Quiropraxia 2010Dokumen38 halamanApostila Quiropraxia 2010gonfinho100% (7)
- Imunologia – ciência experimentalDokumen8 halamanImunologia – ciência experimentalnayaraBelum ada peringkat
- Imuno IIDokumen26 halamanImuno IIJonatan Dos Santos De OliveiraBelum ada peringkat
- Resumo da Imunologia: Introdução à Imunidade Inata e AdquiridaDokumen16 halamanResumo da Imunologia: Introdução à Imunidade Inata e AdquiridaClaudiasenatore SenatoreBelum ada peringkat
- 4 - Imunidade Inata, Inflamação, Citocinas, Migração de Leucocitos e Mecanismos Efetores CelularesDokumen11 halaman4 - Imunidade Inata, Inflamação, Citocinas, Migração de Leucocitos e Mecanismos Efetores Celularesmaria eduarda100% (2)
- Resumo Microbiologia Clinica 120726194949 Phpapp02Dokumen20 halamanResumo Microbiologia Clinica 120726194949 Phpapp02Edirlenesara100% (1)
- Imunidade inata contra fungos e protozoáriosDokumen2 halamanImunidade inata contra fungos e protozoáriosFernanda MotaBelum ada peringkat
- Sistema Imunológico e suas Funções BásicasDokumen5 halamanSistema Imunológico e suas Funções BásicasRhuanLopesBelum ada peringkat
- Hipersensibilidade e os tipos de resposta imune exacerbadaDokumen57 halamanHipersensibilidade e os tipos de resposta imune exacerbadaLucimar FonsecaBelum ada peringkat
- Ciências - Prova 7a sérieDokumen1 halamanCiências - Prova 7a sérieJuAlvesBelum ada peringkat
- Farmacologia - Resumo Do GoodmanDokumen35 halamanFarmacologia - Resumo Do Goodmanyurimusso100% (3)
- Aula 04 - ImunologiaDokumen20 halamanAula 04 - ImunologiaJairlane Garcia100% (1)
- Apostila de Imunologia Clínica 2013Dokumen87 halamanApostila de Imunologia Clínica 2013sergiomolivBelum ada peringkat
- Termo de Anuência Formal para Trabalho em AlturaDokumen1 halamanTermo de Anuência Formal para Trabalho em AlturaRecrutamento Alfa Proteções RecrutamentoBelum ada peringkat
- Mecanismos de defesa e agressãoDokumen17 halamanMecanismos de defesa e agressãoLuana SouzaBelum ada peringkat
- Bônus 2-Ebook ImunidadeDokumen28 halamanBônus 2-Ebook Imunidadeairla rodriguesBelum ada peringkat
- Interações parasita-hospedeiroDokumen5 halamanInterações parasita-hospedeiromaria eduardaBelum ada peringkat
- Bactérias anaeróbias e infecções odontogênicasDokumen52 halamanBactérias anaeróbias e infecções odontogênicasvando2006100% (5)
- Antígenos, Anticorpos e suas EstruturasDokumen37 halamanAntígenos, Anticorpos e suas EstruturasLuizAlbuquerqueJr.Belum ada peringkat
- Imunologia Básica - Sistema Imune e Resposta ImuneDokumen2 halamanImunologia Básica - Sistema Imune e Resposta ImuneMaria Fernanda AguiarBelum ada peringkat
- Fatores Intrínsecos e Extrínsecos Que Interferem No Crescimento Microbiano em AlimentosDokumen7 halamanFatores Intrínsecos e Extrínsecos Que Interferem No Crescimento Microbiano em AlimentosSuzaneemartins100% (1)
- Acidentes CasaDokumen50 halamanAcidentes Casaedson rogeri de abreuBelum ada peringkat
- Hipersensibilidades e doenças autoimunesDokumen69 halamanHipersensibilidades e doenças autoimunesDaniloGomes100% (1)
- Atividade - Inflamação. - PatologiaDokumen5 halamanAtividade - Inflamação. - PatologiaWddCreative Marketing DigitalBelum ada peringkat
- AULA - AntibacterianosDokumen85 halamanAULA - AntibacterianosGian PessattoBelum ada peringkat
- Glomerulonefrite: causas, sintomas e tratamentoDokumen6 halamanGlomerulonefrite: causas, sintomas e tratamentoDaniela LonderoBelum ada peringkat
- Imunidade Inata: Primeira Linha de DefesaDokumen63 halamanImunidade Inata: Primeira Linha de DefesaNina Maria100% (1)
- Distúrbios causados pelas respostas imunes: HipersensibilidadeDokumen9 halamanDistúrbios causados pelas respostas imunes: HipersensibilidadeDaniel Gomes100% (1)
- 2º Prova ImunologiaDokumen2 halaman2º Prova ImunologiaRobson Valverde100% (2)
- Tolerância Imunológica: Conceitos e Mecanismos ChaveDokumen44 halamanTolerância Imunológica: Conceitos e Mecanismos Chavecarolaine santosBelum ada peringkat
- Material Da Aula Todos Os Resumos Pediatria 10 Resumo AulaDokumen28 halamanMaterial Da Aula Todos Os Resumos Pediatria 10 Resumo AulaswryxhgtjxhgBelum ada peringkat
- Artigo de RevisãoDokumen19 halamanArtigo de RevisãoRosafatBelum ada peringkat
- Tipos de receitas médicas e legislação sobre prescrição no BrasilDokumen6 halamanTipos de receitas médicas e legislação sobre prescrição no BrasilNonato Oliveira Carvalho JuniorBelum ada peringkat
- Imunologia dos tumores: antígenos tumoraisDokumen15 halamanImunologia dos tumores: antígenos tumoraisEDUARDOBelum ada peringkat
- Recomendações para As Precauções de IsolamentoDokumen47 halamanRecomendações para As Precauções de IsolamentoTiago CunhaBelum ada peringkat
- Resumo Com Tabela Fungos-MicologiaDokumen3 halamanResumo Com Tabela Fungos-Micologiamaria eduardaBelum ada peringkat
- MENINGITES: CAUSAS, SINTOMAS E TRATAMENTODokumen59 halamanMENINGITES: CAUSAS, SINTOMAS E TRATAMENTOAline MostaroBelum ada peringkat
- Resposta Imune Inata e EspecíficaDokumen54 halamanResposta Imune Inata e EspecíficaFernando Nunomura100% (1)
- Cap 9 Resumo Abbas Imunología BásicaDokumen8 halamanCap 9 Resumo Abbas Imunología BásicaMelissa Falcão100% (1)
- 01-Tipos de ImunidadeDokumen17 halaman01-Tipos de ImunidadeLeonardo Chaves100% (1)
- AEP - Doenças Cardiovasculares - LDADokumen19 halamanAEP - Doenças Cardiovasculares - LDAROSANGELA APARECIDA XIMENEZBelum ada peringkat
- Estudo Dirigido de Imunologia para P1Dokumen2 halamanEstudo Dirigido de Imunologia para P1Vanessa Suely Campos100% (2)
- Questões Imunologia ClínicaDokumen6 halamanQuestões Imunologia Clínicalubeck40Belum ada peringkat
- Aula - Medicamentos AntiparasitáriosDokumen25 halamanAula - Medicamentos AntiparasitáriosTatiane Garcia Do Carmo ValerianoBelum ada peringkat
- Imunologia e seus invasoresDokumen194 halamanImunologia e seus invasoresLuiz Fernando Zoch100% (1)
- Hematologia Básica Parte 1 - Laboratório de Analises ClinicasDokumen18 halamanHematologia Básica Parte 1 - Laboratório de Analises ClinicasP2P3ESTATISTICA CIPTURBelum ada peringkat
- Lesão ReversívelDokumen3 halamanLesão ReversívelNatalia Soares VeigaBelum ada peringkat
- O sistema imunitário e suas defesasDokumen16 halamanO sistema imunitário e suas defesasmalcinacmatiasBelum ada peringkat
- Tipos de Leucócitos - Fábio JoséDokumen6 halamanTipos de Leucócitos - Fábio JoséFabim100% (1)
- Mecanismos de Resistencia Bacteriana Aos Antibióticos TCCDokumen37 halamanMecanismos de Resistencia Bacteriana Aos Antibióticos TCCJuliana Francisco NunesBelum ada peringkat
- Obstrução Nasal No Recém-NascidoDokumen7 halamanObstrução Nasal No Recém-NascidoVicente LanaBelum ada peringkat
- Resumo P1 Química MedicinalDokumen31 halamanResumo P1 Química MedicinalMariana Moreira NunesBelum ada peringkat
- Lei Lucas Na EducaçãoDari EverandLei Lucas Na EducaçãoBelum ada peringkat
- Análise comparativa da composição química e de atividades biológicas de cinco genótipos clonais de Schinus terebinthifolius Raddi (aroeira)Dari EverandAnálise comparativa da composição química e de atividades biológicas de cinco genótipos clonais de Schinus terebinthifolius Raddi (aroeira)Belum ada peringkat
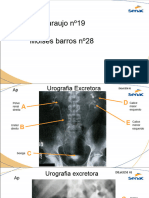
- Arividade Contraste Julio - MoisesDokumen25 halamanArividade Contraste Julio - Moisessfernanda.dutraBelum ada peringkat
- A MagnetoterapiaDokumen7 halamanA MagnetoterapiaMara Carneiro100% (4)
- Lista de Exercícios - BotânicaDokumen1 halamanLista de Exercícios - BotânicaAndreaBarreto100% (1)
- Exercícios Sobre Sist Nervoso Com GabaritoDokumen3 halamanExercícios Sobre Sist Nervoso Com GabaritoNathaniel RossiBelum ada peringkat
- Cavidade Da BocaDokumen11 halamanCavidade Da BocaThalyta DurvalBelum ada peringkat
- Ação Do Ácido Salicílico Na Acne VulgarDokumen174 halamanAção Do Ácido Salicílico Na Acne VulgarAna Paula Pereira CunhaBelum ada peringkat
- Ficha de Avaliação Sumativa 4Dokumen6 halamanFicha de Avaliação Sumativa 4Anabela Rodrigues Gonçalves100% (2)
- 1-Imuno-Hematologia 2009Dokumen88 halaman1-Imuno-Hematologia 2009José Alisson dos SantosBelum ada peringkat
- Metilxantinas em plantasDokumen2 halamanMetilxantinas em plantasLuciana CapetiniBelum ada peringkat
- Pediatria I - Exame Neurológico em CriançasDokumen12 halamanPediatria I - Exame Neurológico em Criançastatchaconti100% (1)
- Hemograma: 02/08/2022 Tutty Isabel Cristina Animale Rs Canoas 11a 7m 1d Macho SRD Canino 616831Dokumen2 halamanHemograma: 02/08/2022 Tutty Isabel Cristina Animale Rs Canoas 11a 7m 1d Macho SRD Canino 616831thariqij3363Belum ada peringkat
- Introdução à Embriologia HumanaDokumen203 halamanIntrodução à Embriologia HumanaBianca TavaresBelum ada peringkat
- Revisão CitoplasmaDokumen3 halamanRevisão CitoplasmaClarice Agudo MenezesBelum ada peringkat
- Trabalho de Anatomia Dia 08 Do 11Dokumen3 halamanTrabalho de Anatomia Dia 08 Do 11Amanda SilvaBelum ada peringkat
- Texto - Imunologia Da Mucosa OralDokumen20 halamanTexto - Imunologia Da Mucosa OralLuana SouzaBelum ada peringkat
- Sistema CirculatórioDokumen7 halamanSistema CirculatórioGheter CostaBelum ada peringkat
- Coração e Vasos Dos Membros PDFDokumen85 halamanCoração e Vasos Dos Membros PDFMarina MachadoBelum ada peringkat
- Fatores de RetençãoDokumen7 halamanFatores de RetençãoFernanda FariaBelum ada peringkat
- Matriz extracelular, tecidos e glândulasDokumen3 halamanMatriz extracelular, tecidos e glândulasgaah100% (1)
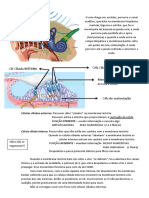
- O som na audição: da membrana timpânica às células ciliadasDokumen2 halamanO som na audição: da membrana timpânica às células ciliadasFonoaudiologiaBelum ada peringkat
- Diretrizes EcocardiografiaDokumen43 halamanDiretrizes EcocardiografiaJuan NatãBelum ada peringkat
- 13 Punção VenosaDokumen56 halaman13 Punção VenosaLetícia Martins100% (1)
- A arte do esquecimentoDokumen8 halamanA arte do esquecimentoRaissa Araújo MendesBelum ada peringkat
- Tabela de HonorariosDokumen5 halamanTabela de HonorariosGeorgia Leandro SalernoBelum ada peringkat