Anda mungkin juga menyukai
- Phenolic Composition, Antioxidant Capacity Andantibacterial Activity of Selected Irish Brassica VegetablesDokumen6 halamanPhenolic Composition, Antioxidant Capacity Andantibacterial Activity of Selected Irish Brassica Vegetablesamit8febBelum ada peringkat
- Evaluation of Benzopyrene in Food From China by High-PerformanceDokumen11 halamanEvaluation of Benzopyrene in Food From China by High-PerformanceJared Nun Barrenechea BacaBelum ada peringkat
- Farhadi An 2010Dokumen5 halamanFarhadi An 2010Anonymous NZHwrtBelum ada peringkat
- Pheniloc Content and Specilay Anosynic ContentDokumen8 halamanPheniloc Content and Specilay Anosynic ContentHarshaka Maduwatha JansBelum ada peringkat
- 1 不同食物PAH含量及风险Dokumen8 halaman1 不同食物PAH含量及风险Mengjiao ZHUBelum ada peringkat
- Synopsis 101 NewDokumen11 halamanSynopsis 101 NewRahul YadavBelum ada peringkat
- Whole Flour and Protein Hydrolysate From Common Beans Reduce The Inflammation in Balb C Mice Fed With High Fat High Cholesterol DietDokumen10 halamanWhole Flour and Protein Hydrolysate From Common Beans Reduce The Inflammation in Balb C Mice Fed With High Fat High Cholesterol DietasdBelum ada peringkat
- Characterization of Bioactive Compounds in Colored Potato (Solanum Tuberosum L.) Cultivars Grown With Conventional, Organic, and Biodynamic MethodsDokumen13 halamanCharacterization of Bioactive Compounds in Colored Potato (Solanum Tuberosum L.) Cultivars Grown With Conventional, Organic, and Biodynamic MethodsShirley EspirituBelum ada peringkat
- Domingo 2016Dokumen7 halamanDomingo 2016charles.lucas008Belum ada peringkat
- The Content of Polyphenols in Coffee Beans As Roasting, Origin and Storage EffectDokumen8 halamanThe Content of Polyphenols in Coffee Beans As Roasting, Origin and Storage EffectBrandon HeatBelum ada peringkat
- Effect of Mixed Starters On Proteolysis and Formation of Biogenic Amines in Dry Fermented Mutton SausagesDokumen15 halamanEffect of Mixed Starters On Proteolysis and Formation of Biogenic Amines in Dry Fermented Mutton SausagesRoberto DegenhardtBelum ada peringkat
- Domestic Cooking Methods Affect The Nutritional Quality of Red CabbageDokumen6 halamanDomestic Cooking Methods Affect The Nutritional Quality of Red CabbageAndrea Osorio AlturoBelum ada peringkat
- Effects of Antimicrobial and Antioxidant Activities of Spice Extracts On Raw Chicken Meat QualityDokumen10 halamanEffects of Antimicrobial and Antioxidant Activities of Spice Extracts On Raw Chicken Meat QualityAmiliaBelum ada peringkat
- Goji Berry (Lycium Barbarum) Composition and Health Effects A Review PDFDokumen10 halamanGoji Berry (Lycium Barbarum) Composition and Health Effects A Review PDFUno MendozaBelum ada peringkat
- 1 s2.0 S0963996917308591 MainDokumen10 halaman1 s2.0 S0963996917308591 MainNatalia BeltranBelum ada peringkat
- Heterocyclic Amines and Polycyclic Aromatic Hydrocarbons in Commercial Ready-To-Eat Meat Products On UK MarketDokumen34 halamanHeterocyclic Amines and Polycyclic Aromatic Hydrocarbons in Commercial Ready-To-Eat Meat Products On UK MarketCarlos LopesBelum ada peringkat
- Analyzing Cranberry Bioactive Compounds: Critical Reviews in Food Science and NutritionDokumen18 halamanAnalyzing Cranberry Bioactive Compounds: Critical Reviews in Food Science and NutritionI DeusBelum ada peringkat
- Science, 20 (1), 947-954. DOI:10.1080/1828051X.2021.1911695.: Physiology of Thyroid GlandsDokumen5 halamanScience, 20 (1), 947-954. DOI:10.1080/1828051X.2021.1911695.: Physiology of Thyroid GlandsWow writersBelum ada peringkat
- Determination of Xanthohumol in Hops, Food Supplements and Beers by HPLCDokumen16 halamanDetermination of Xanthohumol in Hops, Food Supplements and Beers by HPLCMicheli Legemann MonteBelum ada peringkat
- Applsci 13 01011 v2Dokumen11 halamanApplsci 13 01011 v2rajavinugmailcomBelum ada peringkat
- Edible Mushrooms: Chemical Composition and Nutritional ValueDari EverandEdible Mushrooms: Chemical Composition and Nutritional ValuePenilaian: 4 dari 5 bintang4/5 (1)
- Antioxidant Properties of Potatoes: A Review of Carotenoids, Phenolic Compounds, and AnthocyaninsDokumen11 halamanAntioxidant Properties of Potatoes: A Review of Carotenoids, Phenolic Compounds, and AnthocyaninsKlo JosuaBelum ada peringkat
- FTB 54 489Dokumen8 halamanFTB 54 489Nurjihan FaBelum ada peringkat
- Review Article Is Coffee A Functional Food? : Jose G. Do Rea and Teresa Helena M. Da CostaDokumen10 halamanReview Article Is Coffee A Functional Food? : Jose G. Do Rea and Teresa Helena M. Da CostaJean Carlos Torres MoralesBelum ada peringkat
- HPLC Method Determines Total Vitamin B6 in FoodsDokumen13 halamanHPLC Method Determines Total Vitamin B6 in FoodsMoch Alif Zulfikar JamalBelum ada peringkat
- Coccao e BrassicasDokumen11 halamanCoccao e Brassicasclissia croelhasBelum ada peringkat
- Determina-Glucosinolatos InfrarojosDokumen32 halamanDetermina-Glucosinolatos Infrarojoslmedina23039289Belum ada peringkat
- Heavy Metals in Vegetables and Potential Risk ForDokumen7 halamanHeavy Metals in Vegetables and Potential Risk ForYoselinBelum ada peringkat
- Absorption and Metabolism of XanthophyllDokumen2 halamanAbsorption and Metabolism of XanthophyllJames Lord GalangBelum ada peringkat
- Jurnal Hayati BioconversiDokumen13 halamanJurnal Hayati BioconversiririnoorBelum ada peringkat
- Polycyclic Aromatic CompoundsDokumen15 halamanPolycyclic Aromatic CompoundsChouaib OubaaliBelum ada peringkat
- Chapter 2Dokumen6 halamanChapter 2Jessie Jade DurojaBelum ada peringkat
- Composition of Food DyeDokumen16 halamanComposition of Food DyeRona rose HablaBelum ada peringkat
- ConcentracionDokumen7 halamanConcentracionRodrigo Andrés SanabriaBelum ada peringkat
- Accepted Manuscript: Food ChemistryDokumen38 halamanAccepted Manuscript: Food ChemistryLuqman MileBelum ada peringkat
- Marton, Et Al ('10) - The Role of Sprouts in Human Nutrition - A Review (Marked)Dokumen37 halamanMarton, Et Al ('10) - The Role of Sprouts in Human Nutrition - A Review (Marked)aida-69Belum ada peringkat
- Molecules: Roselle Anthocyanins: Antioxidant Properties and Stability To Heat and PHDokumen13 halamanMolecules: Roselle Anthocyanins: Antioxidant Properties and Stability To Heat and PHNguyen Thi Thuy Duong B1606703Belum ada peringkat
- Chemicals in Food: Environments and ContaminantsDokumen18 halamanChemicals in Food: Environments and ContaminantsseemirrBelum ada peringkat
- Levels of organochlorines in organic and conventional cheesesDokumen8 halamanLevels of organochlorines in organic and conventional cheesesBianca AndreeaBelum ada peringkat
- Effect of Natural Extracts Obtained From Food Industry By-Products On Nutritional Quality and Shelf Life of Chicken Nuggets Enriched With Organic ZN and SeDokumen11 halamanEffect of Natural Extracts Obtained From Food Industry By-Products On Nutritional Quality and Shelf Life of Chicken Nuggets Enriched With Organic ZN and SeAliyya SyahminaBelum ada peringkat
- Aspectos de Seguridad en Productos Cárnicos CocidosDokumen11 halamanAspectos de Seguridad en Productos Cárnicos CocidosAndres PerafánBelum ada peringkat
- Folic Acid Content and Antioxidant Activity of Different Types of Beers Available in Hungarian RetailDokumen10 halamanFolic Acid Content and Antioxidant Activity of Different Types of Beers Available in Hungarian RetailAlexandre PimentelBelum ada peringkat
- Emerging Applications of Metabolomics ToDokumen20 halamanEmerging Applications of Metabolomics ToMariela GonzalezBelum ada peringkat
- Biology and Medicine: The Beneficial Effects of Coffee in Human NutritionDokumen5 halamanBiology and Medicine: The Beneficial Effects of Coffee in Human NutritionstevhenBelum ada peringkat
- Kelompok 10 - 1-s2.0-S002364381730004X-mainDokumen4 halamanKelompok 10 - 1-s2.0-S002364381730004X-mainAngelina NitaBelum ada peringkat
- Paket Informasi Perpustakaan Bblitvet No. 10, 2016 PaketinformasibidangtoksikologidanmikologiDokumen18 halamanPaket Informasi Perpustakaan Bblitvet No. 10, 2016 PaketinformasibidangtoksikologidanmikologiUka KahfianaBelum ada peringkat
- Food Chemistry: Yan Zhao, Shuang-Kui Du, Hanxin Wang, Meng CaiDokumen5 halamanFood Chemistry: Yan Zhao, Shuang-Kui Du, Hanxin Wang, Meng CaiValeriu MunteanuBelum ada peringkat
- Cruciferous VegetablesDokumen13 halamanCruciferous Vegetablesapi-271257230Belum ada peringkat
- Food Chemistry: Wieslaw Wiczkowski, Dorota Szawara-Nowak, Joanna TopolskaDokumen9 halamanFood Chemistry: Wieslaw Wiczkowski, Dorota Szawara-Nowak, Joanna TopolskaDaniel Pulido RojasBelum ada peringkat
- Wu 2019Dokumen8 halamanWu 2019An PhanBelum ada peringkat
- Jfca 2017Dokumen7 halamanJfca 2017Bhupinder SinghBelum ada peringkat
- E3sconf Esmgt2021 07005Dokumen6 halamanE3sconf Esmgt2021 07005Muhammad IkhwannasyirBelum ada peringkat
- 1 s2.0 S0023643822009008 MainDokumen7 halaman1 s2.0 S0023643822009008 Mainmemo loveBelum ada peringkat
- Characterization of 12 Capsicum Varieties by Evaluation of Their CarotenoidDokumen9 halamanCharacterization of 12 Capsicum Varieties by Evaluation of Their CarotenoidcarlosBelum ada peringkat
- Detailed Characterization of Plant Based BurgersDokumen9 halamanDetailed Characterization of Plant Based BurgersRhouen RazeBelum ada peringkat
- MEAT SCIENCE EXTRACTS EXTEND SHELF LIFEDokumen10 halamanMEAT SCIENCE EXTRACTS EXTEND SHELF LIFEmmoradi55Belum ada peringkat
- Bisfenol A e Efeitos Na Reproducao e Comportamento 2014Dokumen25 halamanBisfenol A e Efeitos Na Reproducao e Comportamento 2014Priscilla Ferreira Crespo GutierrezBelum ada peringkat
- MORINDA CITRIFOLIA L. (Noni) - A Review of The Scientific Validation For Its Nutritional and Therapeutic PropertiesDokumen15 halamanMORINDA CITRIFOLIA L. (Noni) - A Review of The Scientific Validation For Its Nutritional and Therapeutic PropertiesDiannokaIhzaGanung100% (1)
- A Comprehensive Review of the Health Benefits of Apples and Apple ComponentsDokumen13 halamanA Comprehensive Review of the Health Benefits of Apples and Apple ComponentsAyishaBelum ada peringkat
- PicrolemmaDokumen16 halamanPicrolemmaMarcos SilvaBelum ada peringkat
- Attachment ADokumen52 halamanAttachment Areesty68_unfearyBelum ada peringkat
- DCXR Is A Differentially Expressed Gene in Brain Metastatic Human Breast Cancer.Dokumen10 halamanDCXR Is A Differentially Expressed Gene in Brain Metastatic Human Breast Cancer.reesty68_unfearyBelum ada peringkat
- Pursuant To The State of Nevada Governor's Guidance For COVID-19 Safety Measures, Masks/facial Coverings Are Required at All TimesDokumen7 halamanPursuant To The State of Nevada Governor's Guidance For COVID-19 Safety Measures, Masks/facial Coverings Are Required at All Timesreesty68_unfearyBelum ada peringkat
- Sustainability 12 07064 PDFDokumen17 halamanSustainability 12 07064 PDFreesty68_unfearyBelum ada peringkat
- AE Program Guide Spring 21Dokumen8 halamanAE Program Guide Spring 21reesty68_unfearyBelum ada peringkat
- MENUS ARE DUE Wednesday Jan 20 : To Your Table Meal Subscription MenuDokumen1 halamanMENUS ARE DUE Wednesday Jan 20 : To Your Table Meal Subscription Menureesty68_unfearyBelum ada peringkat
- What is a SacramentDokumen3 halamanWhat is a Sacramentreesty68_unfearyBelum ada peringkat
- JournalUnifiedStyleSheet2007 PDFDokumen4 halamanJournalUnifiedStyleSheet2007 PDFqueencantikBelum ada peringkat
- How To Get Help or File A ComplaintDokumen2 halamanHow To Get Help or File A Complaintreesty68_unfearyBelum ada peringkat
- 20-21 Complaint Policy and ProcedureDokumen3 halaman20-21 Complaint Policy and Procedurereesty68_unfearyBelum ada peringkat
- JournalUnifiedStyleSheet2007 PDFDokumen4 halamanJournalUnifiedStyleSheet2007 PDFqueencantikBelum ada peringkat
- Memorandum Decision Upholds Inventory Search of VehicleDokumen6 halamanMemorandum Decision Upholds Inventory Search of Vehiclereesty68_unfearyBelum ada peringkat
- Most Frontline Staff at California Nursing Homes See Infections at WorkDokumen6 halamanMost Frontline Staff at California Nursing Homes See Infections at Workreesty68_unfearyBelum ada peringkat
- UHN Study With SDM July 17 2020Dokumen3 halamanUHN Study With SDM July 17 2020reesty68_unfearyBelum ada peringkat
- 2020 July 19Dokumen6 halaman2020 July 19reesty68_unfearyBelum ada peringkat
- American Public Health', BDokumen4 halamanAmerican Public Health', Breesty68_unfearyBelum ada peringkat
- US-India SEP Joint Statement - Issued July 17 2020Dokumen5 halamanUS-India SEP Joint Statement - Issued July 17 2020reesty68_unfearyBelum ada peringkat
- Ia 5544Dokumen2 halamanIa 5544reesty68_unfearyBelum ada peringkat
- FullDokumen18 halamanFullreesty68_unfearyBelum ada peringkat
- 2020.07.17.COVID-19 35 DeathsDokumen3 halaman2020.07.17.COVID-19 35 Deathsreesty68_unfearyBelum ada peringkat
- Planning For A Pandemic: Sandia Researchers Used Complexity Science, Modeling To Create Virus Mitigation StrategiesDokumen8 halamanPlanning For A Pandemic: Sandia Researchers Used Complexity Science, Modeling To Create Virus Mitigation Strategiesreesty68_unfearyBelum ada peringkat
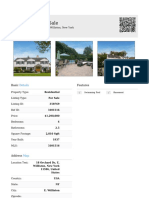
- Residential For Sale: DetailsDokumen2 halamanResidential For Sale: Detailsreesty68_unfearyBelum ada peringkat
- UST PhaseIVPriorities 07.17.20Dokumen3 halamanUST PhaseIVPriorities 07.17.20reesty68_unfearyBelum ada peringkat
- United States Court of Appeals For The Ninth Circuit: Plaintiff-AppelleeDokumen35 halamanUnited States Court of Appeals For The Ninth Circuit: Plaintiff-Appelleereesty68_unfearyBelum ada peringkat
- FREE ROBUX GENERATORDokumen4 halamanFREE ROBUX GENERATORreesty68_unfearyBelum ada peringkat
- Litigator of The Week: How Gibson Dunn Helped Hit Print On Mary Trump's Best-SellerDokumen2 halamanLitigator of The Week: How Gibson Dunn Helped Hit Print On Mary Trump's Best-Sellerreesty68_unfearyBelum ada peringkat
- CP 030919 Fibre EngDokumen2 halamanCP 030919 Fibre Engreesty68_unfearyBelum ada peringkat
- Franco-Nevada Enters Into Strategic Relationship With Continental Resources, IncDokumen2 halamanFranco-Nevada Enters Into Strategic Relationship With Continental Resources, Increesty68_unfearyBelum ada peringkat
- Remote Sensing: A Data-Driven Approach To Classifying Wave Breaking in Infrared ImageryDokumen10 halamanRemote Sensing: A Data-Driven Approach To Classifying Wave Breaking in Infrared Imageryreesty68_unfearyBelum ada peringkat
- IOSA Self Evaluation Form - 31 October 2014Dokumen45 halamanIOSA Self Evaluation Form - 31 October 2014pknight2010Belum ada peringkat
- Schneider - Cptg010 en (Print)Dokumen16 halamanSchneider - Cptg010 en (Print)el_koptan00857693Belum ada peringkat
- Lecture 1 Biochem 232 CellsDokumen13 halamanLecture 1 Biochem 232 CellsaelmowafyBelum ada peringkat
- MPQ2908 - 48V Buck Converter Automotive Reference DesignDokumen4 halamanMPQ2908 - 48V Buck Converter Automotive Reference DesignShubham KaklijBelum ada peringkat
- Risk Management Report SummaryDokumen61 halamanRisk Management Report SummaryIamCcj100% (1)
- Chambal Cable Stayed Bridge Connecting ShoresDokumen6 halamanChambal Cable Stayed Bridge Connecting Shoresafzal taiBelum ada peringkat
- Project Report On Air FilterDokumen5 halamanProject Report On Air FilterEIRI Board of Consultants and PublishersBelum ada peringkat
- 2005-05-12Dokumen18 halaman2005-05-12The University Daily KansanBelum ada peringkat
- NTS - Candidate (Portal)Dokumen1 halamanNTS - Candidate (Portal)Noureen FatimaBelum ada peringkat
- Naskah PublikasiDokumen14 halamanNaskah PublikasiNirmala malaBelum ada peringkat
- Causes of DyspneaDokumen9 halamanCauses of DyspneaHanis Afiqah Violet MeowBelum ada peringkat
- 06wk48 - CSSR Improvement Fach Power Trial Phase II - VDF UKDokumen19 halaman06wk48 - CSSR Improvement Fach Power Trial Phase II - VDF UKsyedBelum ada peringkat
- Flexible and Alternative Seating: in ClassroomsDokumen5 halamanFlexible and Alternative Seating: in ClassroomsweningBelum ada peringkat
- Rundingan Perdagangan Antara Malaysia Dan Indonesia Di Wisma Putra, Kuala Lumpur 1967Dokumen15 halamanRundingan Perdagangan Antara Malaysia Dan Indonesia Di Wisma Putra, Kuala Lumpur 1967nixyingboBelum ada peringkat
- Kuan Yin 100 Divine Lots InterpretationDokumen30 halamanKuan Yin 100 Divine Lots InterpretationEsperanza Theiss100% (2)
- Training and Supervision of Health Care WorkersDokumen12 halamanTraining and Supervision of Health Care WorkerspriyankaBelum ada peringkat
- Chin Cup Therapy An Effective Tool For The Correction of Class III Malocclusion in Mixed and Late Deciduous DentitionsDokumen6 halamanChin Cup Therapy An Effective Tool For The Correction of Class III Malocclusion in Mixed and Late Deciduous Dentitionschic organizerBelum ada peringkat
- Group 17-Solid State NMR - Questions-AnswersDokumen5 halamanGroup 17-Solid State NMR - Questions-AnswersOmkar AkulwarBelum ada peringkat
- El Bill PDFDokumen2 halamanEl Bill PDFvinodBelum ada peringkat
- The Pentester BlueprintDokumen27 halamanThe Pentester Blueprintjames smith100% (1)
- Medical PhysicsDokumen81 halamanMedical Physicsroni roniBelum ada peringkat
- Delta C200 Series AC Drives PDFDokumen5 halamanDelta C200 Series AC Drives PDFspBelum ada peringkat
- Cd6352 Lee Swee LeongDokumen24 halamanCd6352 Lee Swee LeongFrea Marie SarabiaBelum ada peringkat
- MTD CalculationsDokumen50 halamanMTD CalculationsLeny HrBelum ada peringkat
- D Formation Damage StimCADE FDADokumen30 halamanD Formation Damage StimCADE FDAEmmanuel EkwohBelum ada peringkat
- VERALLIA WHITE-BOOK EN March2022 PDFDokumen48 halamanVERALLIA WHITE-BOOK EN March2022 PDFEugenio94Belum ada peringkat
- Regional Ecology Test ScoringDokumen14 halamanRegional Ecology Test Scoringaisyah Wardah201Belum ada peringkat
- Secondary AssessmentsDokumen12 halamanSecondary Assessmentsapi-338389967Belum ada peringkat
- SAP Technical Consultant resumeDokumen11 halamanSAP Technical Consultant resumeKallol BhowmickBelum ada peringkat
- Biosafety FH Guidance Guide Good Manufacturing Practice enDokumen40 halamanBiosafety FH Guidance Guide Good Manufacturing Practice enMaritsa PerHerBelum ada peringkat