Anda mungkin juga menyukai
- Introduction To Enhanced Oil Recovery EOR Processes and Bioremediation of Oil-Contaminated SitesDokumen330 halamanIntroduction To Enhanced Oil Recovery EOR Processes and Bioremediation of Oil-Contaminated SitesIrsani Fenilia100% (1)
- 206 Lab Ex - 3 - Media PreparationDokumen4 halaman206 Lab Ex - 3 - Media PreparationVia SongcalBelum ada peringkat
- Part2MICRO (COMPILED)Dokumen9 halamanPart2MICRO (COMPILED)Thea GonzalesBelum ada peringkat
- InTech-Algal Biorefinery For Biodiesel ProductionDokumen34 halamanInTech-Algal Biorefinery For Biodiesel Productionchelsea_fc43100% (1)
- Carbohydrate CatabolismDokumen4 halamanCarbohydrate CatabolismNguyễn QuangBelum ada peringkat
- 1116 Usp 32 Microbiological Evaluation of Clean Rooms and Other Controlled EnvironmentsDokumen29 halaman1116 Usp 32 Microbiological Evaluation of Clean Rooms and Other Controlled EnvironmentsMauricio Dimaté CamargoBelum ada peringkat
- Final Interim Report Myza 25034Dokumen18 halamanFinal Interim Report Myza 25034hasnizaBelum ada peringkat
- Iso 15213-1-2023Dokumen28 halamanIso 15213-1-2023JOSEPH100% (7)
- 2014 Raed Bacterial Biodegradation of Crude Oil Using LocalDokumen8 halaman2014 Raed Bacterial Biodegradation of Crude Oil Using LocalFaqiangBelum ada peringkat
- A Review of The Mechanism of Microbial Degradation of Petroleum PollutionDokumen5 halamanA Review of The Mechanism of Microbial Degradation of Petroleum PollutionFIORELLA MARYCIELO CIPRIANO RAMOSBelum ada peringkat
- Biodegradation ofDokumen9 halamanBiodegradation ofMaverick CFBelum ada peringkat
- In Situ Biodiesel Production From Residual Oil Recovered From Spent Bleaching EarthDokumen6 halamanIn Situ Biodiesel Production From Residual Oil Recovered From Spent Bleaching EarthAgung SiswahyuBelum ada peringkat
- Manuscript 107501-0606 IJCEE-Biosurfactant Produced by Azotobacter VinelandiiDokumen8 halamanManuscript 107501-0606 IJCEE-Biosurfactant Produced by Azotobacter VinelandiikihelmyBelum ada peringkat
- Production of Lipopeptide Biosurfactants by Bacillus Atrophaeus 5-2a and Their Potential Use in Microbial Enhanced Oil RecoveryDokumen11 halamanProduction of Lipopeptide Biosurfactants by Bacillus Atrophaeus 5-2a and Their Potential Use in Microbial Enhanced Oil Recoverymac1erBelum ada peringkat
- Purwwasena 2009Dokumen8 halamanPurwwasena 2009Emharion DinBelum ada peringkat
- 2016 Al-Bahry Article - PotentialInHeavyOilBiodegradatDokumen13 halaman2016 Al-Bahry Article - PotentialInHeavyOilBiodegradatFaqiangBelum ada peringkat
- This Is A Sample Cover Image For This Issue. The Actual Cover Is Not Yet Available at This Time.Dokumen10 halamanThis Is A Sample Cover Image For This Issue. The Actual Cover Is Not Yet Available at This Time.Christi Gresya Monica PurbaBelum ada peringkat
- Process Simulation and Economic Analysis of Biodiesel Production From Waste Cooking Oil With Membrane BioreactorDokumen8 halamanProcess Simulation and Economic Analysis of Biodiesel Production From Waste Cooking Oil With Membrane Bioreactorعزالدين حسنBelum ada peringkat
- Jurnal PenelitianDokumen10 halamanJurnal PenelitianHelmi BaharBelum ada peringkat
- Hanafy Et Al, 2016Dokumen7 halamanHanafy Et Al, 2016Claudia HerreraBelum ada peringkat
- Microorganisms 12 00318Dokumen14 halamanMicroorganisms 12 00318Gus MalBelum ada peringkat
- Journal of Petroleum Science and Engineering: Jianjie Niu, Qi Liu, Jing LV, Bo PengDokumen11 halamanJournal of Petroleum Science and Engineering: Jianjie Niu, Qi Liu, Jing LV, Bo Pengwahyu suci rahmaniBelum ada peringkat
- Biomass and Bioenergy: Z.N. Akhlisah, R. Yunus, Z.Z. Abidin, B.Y. Lim, D. KaniaDokumen20 halamanBiomass and Bioenergy: Z.N. Akhlisah, R. Yunus, Z.Z. Abidin, B.Y. Lim, D. KaniaPutri sinagaBelum ada peringkat
- Environmental Studies On The Microbial Degradation of Oil Hydrocarbons and Its Application in Lebanese Oil Polluted Coastal and Marine EcosystemDokumen18 halamanEnvironmental Studies On The Microbial Degradation of Oil Hydrocarbons and Its Application in Lebanese Oil Polluted Coastal and Marine EcosystemAgung Raden MangkubumiBelum ada peringkat
- 1 s2.0 S1369703X15001795 MainDokumen11 halaman1 s2.0 S1369703X15001795 MainNaiara KrelingBelum ada peringkat
- Green Heterogeneous Base Catalyst From Ripe and Unripe Plantain Peels Mixture For The Transesterification of Waste Cooking OilDokumen11 halamanGreen Heterogeneous Base Catalyst From Ripe and Unripe Plantain Peels Mixture For The Transesterification of Waste Cooking OilAldo IqbalBelum ada peringkat
- The Effect of Economic Variables On A Bio Refinery For Biodiesel ProductionDokumen14 halamanThe Effect of Economic Variables On A Bio Refinery For Biodiesel ProductionAqsa ChBelum ada peringkat
- I.A Jimoh, Rudyk S.N and Søgaard E.GDokumen28 halamanI.A Jimoh, Rudyk S.N and Søgaard E.Graanja2Belum ada peringkat
- BiodieselDokumen11 halamanBiodieselArnold Murphy Elouma NdingaBelum ada peringkat
- 2016 - Hydroprocessing of Carinata Oil For Hydrocarbon Biofuel OverMo-Zn-Al2O3Dokumen9 halaman2016 - Hydroprocessing of Carinata Oil For Hydrocarbon Biofuel OverMo-Zn-Al2O3dddinzBelum ada peringkat
- Bacterial Community Composition in Produced Water of Diyarbakır Oil Fields in TurkeyDokumen14 halamanBacterial Community Composition in Produced Water of Diyarbakır Oil Fields in TurkeytugceBelum ada peringkat
- Analytical Chemistry Research: Maulidiyah, Muhammad Nurdin, Fetty Fatma, Muh. Natsir, Dwiprayogo WibowoDokumen9 halamanAnalytical Chemistry Research: Maulidiyah, Muhammad Nurdin, Fetty Fatma, Muh. Natsir, Dwiprayogo WibowoOnime No IchinoseBelum ada peringkat
- Investigation of The Activity of Acinetobacter Calcoaceticus Biodemulsifier To Break Stable Water in Oil EmulsionsDokumen7 halamanInvestigation of The Activity of Acinetobacter Calcoaceticus Biodemulsifier To Break Stable Water in Oil EmulsionsMohamed AlhayaniBelum ada peringkat
- Effect of Biodiesel Concentration On Corrosion of Carbon Steel by Serratia MarcescensDokumen7 halamanEffect of Biodiesel Concentration On Corrosion of Carbon Steel by Serratia MarcescensVivi AisahBelum ada peringkat
- Catalyst Acids Ie901175dDokumen4 halamanCatalyst Acids Ie901175dZamri FadilahBelum ada peringkat
- Sdarticle6 With Cover Page v2Dokumen13 halamanSdarticle6 With Cover Page v2Tarun UppalaBelum ada peringkat
- Research ArticleDokumen12 halamanResearch ArticlePranavBelum ada peringkat
- Energy Conversion and ManagementDokumen9 halamanEnergy Conversion and ManagementSriArthiBelum ada peringkat
- Microbial Biotechnology For Enhancing Oil Recovery: Current Developments and Future ProspectsDokumen12 halamanMicrobial Biotechnology For Enhancing Oil Recovery: Current Developments and Future ProspectsEmeka ChinakaBelum ada peringkat
- Aspen HYSYS Simulation For Biodiesel Production FRDokumen7 halamanAspen HYSYS Simulation For Biodiesel Production FRعزالدين حسنBelum ada peringkat
- Catalysts 12 00717 v2Dokumen31 halamanCatalysts 12 00717 v2Kiel JohnBelum ada peringkat
- Energy TechDokumen10 halamanEnergy TechNisa NisaBelum ada peringkat
- Research Article: Process Optimization For Biodiesel Production From Corn Oil and Its Oxidative StabilityDokumen10 halamanResearch Article: Process Optimization For Biodiesel Production From Corn Oil and Its Oxidative StabilityDevina VeriyansariBelum ada peringkat
- Elboulifi 2010Dokumen10 halamanElboulifi 2010Lorraine OliveiraBelum ada peringkat
- R19 Mohebali and Ball 2008Dokumen15 halamanR19 Mohebali and Ball 2008Andres RojasBelum ada peringkat
- Liang Et Al.2015 Springer Protocol (Temp)Dokumen16 halamanLiang Et Al.2015 Springer Protocol (Temp)Robert CoffinBelum ada peringkat
- A Review of Microbial Enhanced Oil RecoveryDokumen12 halamanA Review of Microbial Enhanced Oil Recoverysohil cherkulwarBelum ada peringkat
- 10.1515 - Pac 2017 0703Dokumen10 halaman10.1515 - Pac 2017 0703LCOLimaaaBelum ada peringkat
- Jatropha CurcasDokumen1 halamanJatropha CurcasAhmad YuhardiBelum ada peringkat
- 2Dokumen9 halaman2Diah Wahyu NBelum ada peringkat
- Influence of Free Fatty Acid Content in Biodiesel Production On Non-Edible OilsDokumen6 halamanInfluence of Free Fatty Acid Content in Biodiesel Production On Non-Edible OilsAnton AlgrinovBelum ada peringkat
- Introduction and ObjectivesDokumen7 halamanIntroduction and ObjectivesLale CovetBelum ada peringkat
- Preparation and Characterization of Caso - Sio - Cao/So Composite For Biodiesel ProductionDokumen8 halamanPreparation and Characterization of Caso - Sio - Cao/So Composite For Biodiesel ProductionyohannesBelum ada peringkat
- Gassara Et AlDokumen6 halamanGassara Et Almushtaq521Belum ada peringkat
- The Effects of Catalysts in Biodiesel ProductionDokumen41 halamanThe Effects of Catalysts in Biodiesel ProductionAnonymous VpLeMjIBelum ada peringkat
- Optimization and Heat and Water Integration For Biodiesel Production From Cooking Oil and AlgaeDokumen38 halamanOptimization and Heat and Water Integration For Biodiesel Production From Cooking Oil and AlgaeMaraym FineBelum ada peringkat
- Applied Microbiology: Petroleum and Hydrocarbon MicrobiologyDokumen22 halamanApplied Microbiology: Petroleum and Hydrocarbon MicrobiologyabavoBelum ada peringkat
- IJCHE - Volume 12 - Issue 3 - Pages 55-76Dokumen22 halamanIJCHE - Volume 12 - Issue 3 - Pages 55-76alexanderBelum ada peringkat
- مصدر المطلوبDokumen12 halamanمصدر المطلوبaliBelum ada peringkat
- Biodiesel Production From Waste Cooking Oil in A Magnetically Fluidized Bed Reactor Using Whole-Cell BiocatalystsDokumen9 halamanBiodiesel Production From Waste Cooking Oil in A Magnetically Fluidized Bed Reactor Using Whole-Cell BiocatalystsFrancisco André Mayorga YuntulBelum ada peringkat
- Preparation, Characterization and Application of Zeolite-Based CatalystDokumen6 halamanPreparation, Characterization and Application of Zeolite-Based CatalystIlham Kiki ShahilaBelum ada peringkat
- Biocatalytic Desulfurization (BDS) of Petrodiesel Fuels: ReviewDokumen15 halamanBiocatalytic Desulfurization (BDS) of Petrodiesel Fuels: ReviewOctavianus RudyBelum ada peringkat
- El-Naas Et Al., (2013) PRWWDokumen7 halamanEl-Naas Et Al., (2013) PRWWAndrea Carolina Cubillos OnateBelum ada peringkat
- Petroleo BacteriasDokumen6 halamanPetroleo BacteriasGABRIELA YELIXA SULLON PURIZACABelum ada peringkat
- Advances in Biofeedstocks and Biofuels, Volume 2: Production Technologies for BiofuelsDari EverandAdvances in Biofeedstocks and Biofuels, Volume 2: Production Technologies for BiofuelsLalit Kumar SinghBelum ada peringkat
- G16 Virtual Lab 2 Bacteria Growth Virtual Lab 1Dokumen9 halamanG16 Virtual Lab 2 Bacteria Growth Virtual Lab 1Isabel Maxine DavidBelum ada peringkat
- A Critical Review of Measurement Uncertainty in The Enumeration of Food Micro-Organisms - Pdf-Cdekey - DSLKOK4FQDJRD7OZ2ZGTG6FNJSRZAZ24Dokumen24 halamanA Critical Review of Measurement Uncertainty in The Enumeration of Food Micro-Organisms - Pdf-Cdekey - DSLKOK4FQDJRD7OZ2ZGTG6FNJSRZAZ24Carolina MontoyaBelum ada peringkat
- Is 5402 2012 PDFDokumen17 halamanIs 5402 2012 PDFakBelum ada peringkat
- PseudomonasDokumen9 halamanPseudomonasMaria Alejandra Nuñez CarvajalBelum ada peringkat
- Approaching Microbiological Method Validation IVTDokumen18 halamanApproaching Microbiological Method Validation IVTPiruzi MaghlakelidzeBelum ada peringkat
- Media Preparation For Plant Tissue Culture: MethodDokumen5 halamanMedia Preparation For Plant Tissue Culture: MethodNguyenDungBelum ada peringkat
- Brevetto CN102823429A - Novel Morel Cultivation Method - Google BrevettiDokumen6 halamanBrevetto CN102823429A - Novel Morel Cultivation Method - Google BrevettinicungBelum ada peringkat
- Handbook 6th Sem PharmacyDokumen35 halamanHandbook 6th Sem PharmacyJai MinhasBelum ada peringkat
- M1245Dokumen2 halamanM1245Chairil AnwarBelum ada peringkat
- SabouraudDokumen3 halamanSabouraudLesly ReyesBelum ada peringkat
- Mfhpb21-Eng - Enumeration of Staphylococcus Aureus in FoodsDokumen14 halamanMfhpb21-Eng - Enumeration of Staphylococcus Aureus in FoodsPedro Carrillo PeredoBelum ada peringkat
- Animal Cell CultureDokumen8 halamanAnimal Cell CulturetharsinibalayogeshBelum ada peringkat
- MYP Agar: Experimental Procedure and EvaluationDokumen2 halamanMYP Agar: Experimental Procedure and EvaluationhechunBelum ada peringkat
- Chemical Compositions of Banana Peels (Musa Sapientum) Fruits Cultivated in Malaysia Using Proximate AnalysisDokumen6 halamanChemical Compositions of Banana Peels (Musa Sapientum) Fruits Cultivated in Malaysia Using Proximate AnalysiszawBelum ada peringkat
- MIBO-111 PracticalDokumen55 halamanMIBO-111 PracticalShinchan DoremonBelum ada peringkat
- Difco & BBL Manual: Manual of Microbiological Culture MediaDokumen2 halamanDifco & BBL Manual: Manual of Microbiological Culture MediaYacine SarniBelum ada peringkat
- 03.0784 Micro Prod BrochureDokumen10 halaman03.0784 Micro Prod BrochureHassan HoudoudBelum ada peringkat
- Module 3 (Updated) MicrobiologyDokumen6 halamanModule 3 (Updated) MicrobiologyJohn Mark Gallano CanayonBelum ada peringkat
- Nutrient Agar: Read More About How To Make Nutrient Agar at HomeDokumen2 halamanNutrient Agar: Read More About How To Make Nutrient Agar at HomeBebi WakaBelum ada peringkat
- Cells Culture Counting, Dilution and Plating CalculatorDokumen1 halamanCells Culture Counting, Dilution and Plating CalculatoritaimoBelum ada peringkat
- Decolorization of Polymeric Dyes by A Novel Penicillium IsolateDokumen7 halamanDecolorization of Polymeric Dyes by A Novel Penicillium IsolatecacaBelum ada peringkat
- Bio IADokumen17 halamanBio IALisaBelum ada peringkat
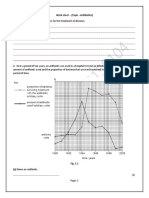
- Work Sheet - (Topic - Antibiotics)Dokumen4 halamanWork Sheet - (Topic - Antibiotics)Jotaro kujoBelum ada peringkat
- Histopathology QuestionsDokumen5 halamanHistopathology QuestionsRey AlegrosoBelum ada peringkat
- Evaluating Performance of 0.1mic FilterDokumen1 halamanEvaluating Performance of 0.1mic FilterAbhijit GhoshBelum ada peringkat