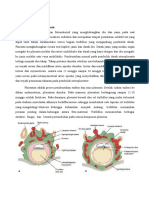
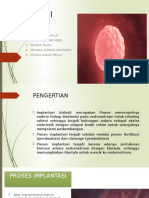
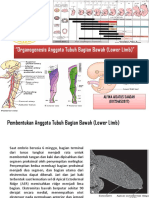
Anda mungkin juga menyukai
- Mikrobiologi Medis I: Patogen dan Mikrobioma ManusiaDari EverandMikrobiologi Medis I: Patogen dan Mikrobioma ManusiaPenilaian: 4 dari 5 bintang4/5 (11)
- Mikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaDari EverandMikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaPenilaian: 2.5 dari 5 bintang2.5/5 (2)
- Fisiologi PlasentaDokumen20 halamanFisiologi PlasentaNoor Widyani29% (7)
- Askeb KomunitasDokumen66 halamanAskeb Komunitasdedy setyawan67% (6)
- Proses EMBRIOGENESISDokumen8 halamanProses EMBRIOGENESIStasyadelizaBelum ada peringkat
- Embriologi Kedokteran LangmanDokumen7 halamanEmbriologi Kedokteran LangmanMuslihudin ahmadBelum ada peringkat
- Makalah PlasentaDokumen23 halamanMakalah PlasentaYesica Septianty33% (3)
- Ekstraseluler MatriksDokumen4 halamanEkstraseluler MatriksWaode Rifa Adhiani67% (3)
- Anatomi, Fisiologi Plasenta, Dan Cairan Amnion (Pres)Dokumen28 halamanAnatomi, Fisiologi Plasenta, Dan Cairan Amnion (Pres)dnazaryBelum ada peringkat
- Kelainan Plasenta Dan Tali PusatDokumen36 halamanKelainan Plasenta Dan Tali PusatDevi Syam100% (1)
- Embriologi Umum PDFDokumen16 halamanEmbriologi Umum PDFDisi Muhaymin RBelum ada peringkat
- Fisiologi PlasentaDokumen9 halamanFisiologi PlasentaFebrinata Mahadika100% (1)
- Transisi Demografi Dan EpidemiologiDokumen12 halamanTransisi Demografi Dan Epidemiologidedy setyawanBelum ada peringkat
- Perkembangan Embrio ManusiaDokumen15 halamanPerkembangan Embrio ManusiaNovi HariyaniBelum ada peringkat
- 3 Fisio PlasentaDokumen17 halaman3 Fisio PlasentaEmir FakhrudinBelum ada peringkat
- Anfis KehamilanDokumen59 halamanAnfis KehamilanZelly Bakri100% (1)
- NidasiDokumen11 halamanNidasiFebyami SiburianBelum ada peringkat
- Proses Pembentukan PlasentaDokumen5 halamanProses Pembentukan PlasentaRiffinola Mepa Venesia IIBelum ada peringkat
- Perkemb - PlasentaDokumen34 halamanPerkemb - PlasentaNita AndiniBelum ada peringkat
- ImplantasiiiiiklpDokumen16 halamanImplantasiiiiiklpSakina Ayu PutriBelum ada peringkat
- Cakram Mudigah BilaminarDokumen5 halamanCakram Mudigah BilaminarEviBelum ada peringkat
- Embrioblast Kelompok 4Dokumen11 halamanEmbrioblast Kelompok 4QurotulAqyunBelum ada peringkat
- Icha SRP - Makalah Fertilisasi Dan Gastrulasi1323Dokumen8 halamanIcha SRP - Makalah Fertilisasi Dan Gastrulasi1323راسیلاواتیBelum ada peringkat
- PLASENTADokumen3 halamanPLASENTAwita zaharaBelum ada peringkat
- (Fertilisasi-Gastrulasi) Puan Nurul RamadhaniDokumen10 halaman(Fertilisasi-Gastrulasi) Puan Nurul RamadhaniراسیلاواتیBelum ada peringkat
- IMPLANTASI (Adnan, UNM)Dokumen32 halamanIMPLANTASI (Adnan, UNM)Adnan Gassing100% (5)
- SK 2 Blok Repro KehamilanDokumen40 halamanSK 2 Blok Repro KehamilanUlfaTitiswariBelum ada peringkat
- Fisiologi PLASENTADokumen17 halamanFisiologi PLASENTAFebrinata MahadikaBelum ada peringkat
- Kel 7. Implantasi Dan Perkembangan PlasentaDokumen27 halamanKel 7. Implantasi Dan Perkembangan PlasentaPutriBelum ada peringkat
- Implantasi Dan PlasentaDokumen16 halamanImplantasi Dan PlasentaMaditaningtyas Hawwa ZuwandaBelum ada peringkat
- PERKEMBANGAN PLASENTADokumen3 halamanPERKEMBANGAN PLASENTALalu Anzuru Wahiyal QamaruBelum ada peringkat
- PLASENTA DAN PERKEMBANGAN SELAPUT JANINDokumen37 halamanPLASENTA DAN PERKEMBANGAN SELAPUT JANINAde Nurhasanah AmirBelum ada peringkat
- Fisiologi PlasentaDokumen11 halamanFisiologi PlasentaFara SippiriliBelum ada peringkat
- ObgynDokumen33 halamanObgynmuhamad deniansyahBelum ada peringkat
- Bu Rila Embriologi KLMPK 3Dokumen19 halamanBu Rila Embriologi KLMPK 3Siti TaifahBelum ada peringkat
- Fisiologi PlasentaDokumen12 halamanFisiologi PlasentaRanvabesa Cakra Laksa Cita100% (1)
- Makalah Implantasi Dan Plasenta 4Dokumen36 halamanMakalah Implantasi Dan Plasenta 4AfntssybhBelum ada peringkat
- PLASENTADokumen5 halamanPLASENTAasti nurisBelum ada peringkat
- Implantasi Dan Perkembangan PlasentaDokumen4 halamanImplantasi Dan Perkembangan PlasentaFebrinata MahadikaBelum ada peringkat
- Anatomi Fisiologi KehamilanDokumen68 halamanAnatomi Fisiologi KehamilanEndyBelum ada peringkat
- PERTEMUAN 3. Cakram Bilaminer N Trilaminer LiliDokumen12 halamanPERTEMUAN 3. Cakram Bilaminer N Trilaminer LiliRaihana FauzhyaBelum ada peringkat
- Makalah NIDASIDokumen14 halamanMakalah NIDASINjet Chamypar L-w'aeis100% (1)
- Implantasi IiiDokumen31 halamanImplantasi IiiReynaldiSanjayaBelum ada peringkat
- Dasar Teori EmbriogenesisDokumen4 halamanDasar Teori EmbriogenesisDei Unzila RahmahBelum ada peringkat
- OBSTETRI FISIOLOGIDokumen19 halamanOBSTETRI FISIOLOGIRisma PertiwiBelum ada peringkat
- PERKEMBANGAN JANIN DAN STRUKTUR PLASENTADokumen67 halamanPERKEMBANGAN JANIN DAN STRUKTUR PLASENTANurhadijah Nst100% (5)
- Diskus Germ Bilaminar Dan TrilaminarDokumen18 halamanDiskus Germ Bilaminar Dan Trilaminarancha100% (1)
- BLASTOKIST DAN IMPLANTASIDokumen3 halamanBLASTOKIST DAN IMPLANTASIEunHa KimBelum ada peringkat
- Solutio PlasentaDokumen17 halamanSolutio PlasentaRaysha RamadhaniBelum ada peringkat
- Proses NidasiDokumen4 halamanProses NidasiNadia Sani amaliaBelum ada peringkat
- Plasenta, Tali Pusat, Selaput Janin Dan Cairan AmnionDokumen7 halamanPlasenta, Tali Pusat, Selaput Janin Dan Cairan AmnionNaldi SufriBelum ada peringkat
- GastrulaDokumen6 halamanGastruladhio putra100% (1)
- IMPLANTASIDokumen11 halamanIMPLANTASIRahmawaty DukalangBelum ada peringkat
- NIDASIDokumen40 halamanNIDASISiwi TrimulyaniBelum ada peringkat
- PROSES NIDASIDokumen3 halamanPROSES NIDASIfirda0% (1)
- Embriologi UmumDokumen27 halamanEmbriologi UmumRendi Siregar0% (2)
- Anatomi, Fisiologi Plasenta Dan Cairan AmnionDokumen28 halamanAnatomi, Fisiologi Plasenta Dan Cairan Amnionicha100% (1)
- Proses ImplantasiDokumen13 halamanProses ImplantasiollyBelum ada peringkat
- Peran Trofoblas Dalam Perkembangan PlasentaDokumen23 halamanPeran Trofoblas Dalam Perkembangan PlasentanadiaBelum ada peringkat
- Soal SoalDokumen8 halamanSoal Soaldedy setyawanBelum ada peringkat
- Alfina Aisatus SaadahDokumen6 halamanAlfina Aisatus Saadahalfina aisatus saadahBelum ada peringkat
- MKDokumen5 halamanMKdedy setyawanBelum ada peringkat
- SAMPUL Epidemiologi DR BudiDokumen3 halamanSAMPUL Epidemiologi DR Budidedy setyawanBelum ada peringkat
- Kelompok 4 Transisi Demografi EpidemioloDokumen21 halamanKelompok 4 Transisi Demografi EpidemioloAldo AshuriBelum ada peringkat
- Filsafat Dan Etik S2 PurwoDokumen1 halamanFilsafat Dan Etik S2 Purwodedy setyawanBelum ada peringkat
- MateriDokumen13 halamanMateridedy setyawanBelum ada peringkat
- Radioterapi SCR Umum TRHDP Infertilitas TranslateDokumen21 halamanRadioterapi SCR Umum TRHDP Infertilitas Translatededy setyawanBelum ada peringkat
- Bab I Pendahuluan 1.1 Latar BelakangDokumen31 halamanBab I Pendahuluan 1.1 Latar Belakangdedy setyawanBelum ada peringkat
- Komplikasi Sindroma Hiperstimulasi OvariumDokumen14 halamanKomplikasi Sindroma Hiperstimulasi OvariumNhoeExanisahBelum ada peringkat
- TRANSISI DEMOGRAFI DI INDONESIADokumen22 halamanTRANSISI DEMOGRAFI DI INDONESIAdedy setyawanBelum ada peringkat
- OrisinalitasDokumen1 halamanOrisinalitasdedy setyawanBelum ada peringkat
- Bab 2Dokumen15 halamanBab 2dedy setyawanBelum ada peringkat
- MateriDokumen12 halamanMateridedy setyawanBelum ada peringkat
- Pengetahuan Dan Ilmu Pengetahuan-1Dokumen46 halamanPengetahuan Dan Ilmu Pengetahuan-1dedy setyawanBelum ada peringkat
- Vaskularisasi Lower LimbDokumen6 halamanVaskularisasi Lower Limbdedy setyawanBelum ada peringkat
- Nama ArwahDokumen1 halamanNama Arwahdedy setyawanBelum ada peringkat
- Bab 1Dokumen31 halamanBab 1dedy setyawanBelum ada peringkat
- CoverDokumen3 halamanCoverdedy setyawanBelum ada peringkat
- Disfungsi EreksiDokumen17 halamanDisfungsi EreksiCicilia GorretiBelum ada peringkat
- Bufas Dan KBDokumen2 halamanBufas Dan KBdedy setyawanBelum ada peringkat
- MateriDokumen12 halamanMateridedy setyawanBelum ada peringkat
- Rencana Anggaran Dana PenelitianDokumen1 halamanRencana Anggaran Dana Penelitiandedy setyawanBelum ada peringkat
- Daftar PustakaDokumen3 halamanDaftar Pustakadedy setyawanBelum ada peringkat
- MateriDokumen12 halamanMateridedy setyawanBelum ada peringkat
- Male Sexual DysfunctionDokumen13 halamanMale Sexual Dysfunctiondedy setyawanBelum ada peringkat