Anda mungkin juga menyukai
- Caso Clínico BlindagemDokumen11 halamanCaso Clínico BlindagemArmando JúniorBelum ada peringkat
- Caso Clínico BlindagemDokumen11 halamanCaso Clínico BlindagemArmando JúniorBelum ada peringkat
- Cirurgia Odontológica APSDokumen8 halamanCirurgia Odontológica APSArmando JúniorBelum ada peringkat
- Caderno Do DentistaDokumen49 halamanCaderno Do DentistaClaudiony Azevêdo100% (2)
- Perio teste conhecimentoDokumen4 halamanPerio teste conhecimentoArmando JúniorBelum ada peringkat
- Técnicas anestésicas para maxila e mandíbulaDokumen21 halamanTécnicas anestésicas para maxila e mandíbulaCHARLES DOS SANTOS NEVES100% (2)
- Apostila Módulo02Dokumen49 halamanApostila Módulo02Armando JúniorBelum ada peringkat
- Manual de Odontologia HospitalarDokumen32 halamanManual de Odontologia HospitalarArmando JúniorBelum ada peringkat
- Apostila Módulo03Dokumen40 halamanApostila Módulo03Armando JúniorBelum ada peringkat
- Guia de endodontia laboratorialDokumen31 halamanGuia de endodontia laboratorialArmando JúniorBelum ada peringkat
- Princípios e estrutura do SUSDokumen22 halamanPrincípios e estrutura do SUSArmando Júnior100% (10)
- Versao Gratis - Apostila SUS para Concursos 2017Dokumen73 halamanVersao Gratis - Apostila SUS para Concursos 2017Natiele Crepaldi100% (1)
- Acessibilida Na Usp de BauruDokumen13 halamanAcessibilida Na Usp de BauruArmando JúniorBelum ada peringkat
- Manifestações orais em pacientes oncológicos tratados com quimioterapiaDokumen1 halamanManifestações orais em pacientes oncológicos tratados com quimioterapiaArmando JúniorBelum ada peringkat
- Apostila EndodontiaDokumen101 halamanApostila EndodontiaArmando JúniorBelum ada peringkat
- Medresumos Imunologia - MHC e Apresentação AntigênicaDokumen5 halamanMedresumos Imunologia - MHC e Apresentação AntigênicaArmando JúniorBelum ada peringkat
- Preparos e restaurações odontológicasDokumen106 halamanPreparos e restaurações odontológicasmarc10oceanbluestarBelum ada peringkat
- O Posto de Trabalho Odontologico - ErgonomiaDokumen10 halamanO Posto de Trabalho Odontologico - ErgonomiaArmando JúniorBelum ada peringkat
- Órgãos Do Sistema ImuneDokumen5 halamanÓrgãos Do Sistema ImuneArmando JúniorBelum ada peringkat
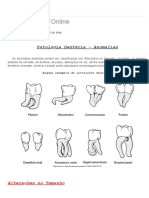
- Anomalias dentárias: microdontia, macrodontia e outras alteraçõesDokumen15 halamanAnomalias dentárias: microdontia, macrodontia e outras alteraçõesArmando JúniorBelum ada peringkat
- Agenesias DentariasDokumen3 halamanAgenesias DentariasArmando JúniorBelum ada peringkat
- Cariologia BásicaDokumen20 halamanCariologia BásicaArmando JúniorBelum ada peringkat
- Farmacologia - Resumo CompletoDokumen95 halamanFarmacologia - Resumo CompletoCarol Pimenta100% (1)
- MEDRESUMOS 2016 - IMUNOLOGIA 08 - MHC e Apresentação Antigênica PDFDokumen8 halamanMEDRESUMOS 2016 - IMUNOLOGIA 08 - MHC e Apresentação Antigênica PDFArmando JúniorBelum ada peringkat
- Posições e movimentos da ATMDokumen9 halamanPosições e movimentos da ATMJoana MartinsBelum ada peringkat
- O Segredo de Um Bom NegocioDokumen2 halamanO Segredo de Um Bom NegocioArmando JúniorBelum ada peringkat
- Estudo Dirigido - BioestatisticaDokumen3 halamanEstudo Dirigido - BioestatisticaArmando JúniorBelum ada peringkat
- Cabelo Caindo MuitoDokumen11 halamanCabelo Caindo MuitoJose Divino0% (1)
- Instruções de Uso Cardioversor - Cardioserv PDFDokumen101 halamanInstruções de Uso Cardioversor - Cardioserv PDFDiogenes Borowsky0% (1)
- Edital 001 2019 Seletivo FMS 2019Dokumen54 halamanEdital 001 2019 Seletivo FMS 2019DAVID MOURABelum ada peringkat
- Parteiras e o Cuidado Com o Nascimento.Dokumen5 halamanParteiras e o Cuidado Com o Nascimento.ZezinhoLucioSilvaBelum ada peringkat
- A Importância Da Pulsão Como Um Dos Conceitos Fundamentais Da Psicanálise.Dokumen19 halamanA Importância Da Pulsão Como Um Dos Conceitos Fundamentais Da Psicanálise.Douglas Naegele100% (2)
- Unimed Nacional PmeDokumen14 halamanUnimed Nacional PmeAlexandre SáBelum ada peringkat
- Teste Avaliação Biologia Geologia Critérios Correção CotaçõesDokumen2 halamanTeste Avaliação Biologia Geologia Critérios Correção CotaçõesMiguelBelum ada peringkat
- Cirurgias ginecológicas e distopias genitaisDokumen23 halamanCirurgias ginecológicas e distopias genitaisAna Carolina Souza TorresBelum ada peringkat
- Estudo malformações embrionáriasDokumen28 halamanEstudo malformações embrionáriasSamy PontesBelum ada peringkat
- Exame Macroscópico e MicroscópicoDokumen4 halamanExame Macroscópico e MicroscópicoEliani Lopes100% (1)
- Domínio Emocional PDFDokumen44 halamanDomínio Emocional PDFDilma Coelho100% (4)
- Os Benefícios Dos Ions NegativosDokumen20 halamanOs Benefícios Dos Ions NegativosRoney KumpelBelum ada peringkat
- Método SocráticoDokumen52 halamanMétodo Socráticopinus_ellioti100% (2)
- Aula 2 - RTM - Bases Da Terapia ManualDokumen6 halamanAula 2 - RTM - Bases Da Terapia ManualPedro MouraBelum ada peringkat
- Revista Alergia ImunopatologiaDokumen21 halamanRevista Alergia ImunopatologiaAnnie OliveiraBelum ada peringkat
- ToxicologiaDokumen68 halamanToxicologiaDiego AndradeBelum ada peringkat
- Cartilha de Orientações Paralisia CerebralDokumen10 halamanCartilha de Orientações Paralisia CerebralMyara CristinyBelum ada peringkat
- Aula 1 Atividade Fisica para Grupos EspeciaisDokumen12 halamanAula 1 Atividade Fisica para Grupos EspeciaisTalita Oliveira50% (2)
- Bula Azitromicina 10135 1181Dokumen2 halamanBula Azitromicina 10135 1181aBelum ada peringkat
- A Alegria Delicada Dos Dias Comuns PDFDokumen65 halamanA Alegria Delicada Dos Dias Comuns PDFMaraCoradelloBelum ada peringkat
- Fisio TonDokumen4 halamanFisio TonGlaucia Simone Oliveira Dos SantosBelum ada peringkat
- TCC - Cannabis Medicinal - Rodrigo Correa Da SilvaDokumen14 halamanTCC - Cannabis Medicinal - Rodrigo Correa Da SilvaRodrigo MovendoBelum ada peringkat
- Introdução A PneumologiaDokumen84 halamanIntrodução A PneumologiamathausmmsBelum ada peringkat
- ISRS: mecanismo de ação e propriedades da sertralina e fluoxetinaDokumen29 halamanISRS: mecanismo de ação e propriedades da sertralina e fluoxetinaRiccardo CasarottiBelum ada peringkat
- Cuidados básicos de higiene em crianças e jovensDokumen2 halamanCuidados básicos de higiene em crianças e jovensAnabela100% (1)
- Manejo da transferência na clínica de adolescentesDokumen32 halamanManejo da transferência na clínica de adolescentesBergsonPaulinoBelum ada peringkat
- Resolucão Normativa - RN Nº - 262, de 1º - de Agosto de 2011Dokumen6 halamanResolucão Normativa - RN Nº - 262, de 1º - de Agosto de 2011Jan JaneBelum ada peringkat
- Proposta de Redação Farias Brito Tema Juventude e AlcoolDokumen1 halamanProposta de Redação Farias Brito Tema Juventude e AlcoolsimoneBelum ada peringkat
- Simulado de Raciocínio ClínicoDokumen3 halamanSimulado de Raciocínio ClínicoJoseph Andrade NeryBelum ada peringkat
- Relatório de estágio em reprodução animalDokumen73 halamanRelatório de estágio em reprodução animalDébora MonteiroBelum ada peringkat