Anda mungkin juga menyukai
- tmpEA98 TMPDokumen2 halamantmpEA98 TMPFrontiersBelum ada peringkat
- Breathing Coordinates Cortico-Hippocampal Dynamics in Mice During of Ine StatesDokumen20 halamanBreathing Coordinates Cortico-Hippocampal Dynamics in Mice During of Ine StatesKris MBelum ada peringkat
- (14796805 - Journal of Endocrinology) The Role of Corticotropin-Releasing Factor in Depression and Anxiety DisordersDokumen12 halaman(14796805 - Journal of Endocrinology) The Role of Corticotropin-Releasing Factor in Depression and Anxiety Disorderskaren Daniela Puma FarinangoBelum ada peringkat
- Differences in Spontaneous and Evoked Neurotransmission ActivitiesDokumen25 halamanDifferences in Spontaneous and Evoked Neurotransmission ActivitiesChrisantus OkakaBelum ada peringkat
- Kuo Etal 2020Dokumen27 halamanKuo Etal 2020林瑜璿Belum ada peringkat
- Commonly Used Meridian PointsDokumen68 halamanCommonly Used Meridian Pointsrcristina762635Belum ada peringkat
- Midbrain Serotonergic Neurons Are Central PH Chemoreceptors: Brief CommunicationsDokumen2 halamanMidbrain Serotonergic Neurons Are Central PH Chemoreceptors: Brief CommunicationsRehaan FayazBelum ada peringkat
- Pone 0088003Dokumen16 halamanPone 0088003jalewa.jaishreeBelum ada peringkat
- Beymer 2016Dokumen11 halamanBeymer 2016Canuto MunozBelum ada peringkat
- OrlDokumen14 halamanOrlchadi_lbBelum ada peringkat
- TMP F204Dokumen2 halamanTMP F204FrontiersBelum ada peringkat
- Invertebrate Hormones: Tissue HormonesDari EverandInvertebrate Hormones: Tissue HormonesU.S. Von EulerBelum ada peringkat
- GSL Marc MalenkaDokumen10 halamanGSL Marc MalenkaDebanjan GoswamiBelum ada peringkat
- Levy and Wagner, 2011Dokumen23 halamanLevy and Wagner, 2011Fati NovBelum ada peringkat
- 4 PhysiologyDokumen16 halaman4 PhysiologyAdel HamadaBelum ada peringkat
- European Journal of Endocrinology) Corticotropin-Releasing Hormone PhysiologyDokumen6 halamanEuropean Journal of Endocrinology) Corticotropin-Releasing Hormone PhysiologyBenedictus Yudha BaskaraBelum ada peringkat
- The Regulation of Sleep and Wakefulness by The Hypothalamic Neuropeptide Orexin/HypocretinDokumen8 halamanThe Regulation of Sleep and Wakefulness by The Hypothalamic Neuropeptide Orexin/HypocretinmeiutaBelum ada peringkat
- Convergence of Excitatory and InhibitoryDokumen21 halamanConvergence of Excitatory and InhibitoryAgus WiyonoBelum ada peringkat
- 76 Prolactin5 PDFDokumen12 halaman76 Prolactin5 PDFSalsabilla SaharaBelum ada peringkat
- LRP1 is a neuronal receptor for α-synuclein uptake and spreadDokumen14 halamanLRP1 is a neuronal receptor for α-synuclein uptake and spreadLilianneBelum ada peringkat
- Nociceptive Stimuli Activate The Hypothalamus-HabeDokumen24 halamanNociceptive Stimuli Activate The Hypothalamus-HabeVissente TapiaBelum ada peringkat
- The Evolution of Emotion and Cognition in the Anterior Cingulate CortexDokumen11 halamanThe Evolution of Emotion and Cognition in the Anterior Cingulate CortexJuan CarcaustoBelum ada peringkat
- Adiccion y Arousal PDFDokumen4 halamanAdiccion y Arousal PDFcalfupanBelum ada peringkat
- The Hypocretins: Setting The Arousal ThresholdDokumen11 halamanThe Hypocretins: Setting The Arousal ThresholdQiantuBelum ada peringkat
- A Minimal Physiologically Based Model of The HPA Axis Under Influence of The Sleep Wake CyclesDokumen9 halamanA Minimal Physiologically Based Model of The HPA Axis Under Influence of The Sleep Wake CyclesPrakash N KBelum ada peringkat
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967Dari EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterBelum ada peringkat
- Learningmemory PDFDokumen8 halamanLearningmemory PDFidjacobsBelum ada peringkat
- Modelling and Analysis of Local FieldDokumen16 halamanModelling and Analysis of Local FieldRomina RoroBelum ada peringkat
- Nihms 710720Dokumen32 halamanNihms 710720Estigma Universidad Del RosarioBelum ada peringkat
- Lateral Hypothalamic Neuropeptides in Reward and Drug AddictionDokumen10 halamanLateral Hypothalamic Neuropeptides in Reward and Drug AddictionCansu UnsalBelum ada peringkat
- Sleep and Brain ActivityDari EverandSleep and Brain ActivityMarcos G. FrankBelum ada peringkat
- Human Pineal Physiology and Functional Signi Wcance of MelatoninDokumen19 halamanHuman Pineal Physiology and Functional Signi Wcance of MelatonincrisbourBelum ada peringkat
- Husos de SueñoDokumen37 halamanHusos de SueñoYair FloresBelum ada peringkat
- The Peripheral Cerebrospinal Fluid Outflow Pathway  " Physiology and Pathophysiology of CSF Recirculation - A Review and HypothesisDokumen16 halamanThe Peripheral Cerebrospinal Fluid Outflow Pathway  " Physiology and Pathophysiology of CSF Recirculation - A Review and Hypothesisمحمد رؤوف خير الدينBelum ada peringkat
- Dampney 2009Dokumen1 halamanDampney 2009elbueno21Belum ada peringkat
- Ni Hms 284366Dokumen27 halamanNi Hms 284366Proyectos SPABelum ada peringkat
- The Neuroendocrine Control of Adaptation: International Series of Monographs in Pure and Applied BiologyDari EverandThe Neuroendocrine Control of Adaptation: International Series of Monographs in Pure and Applied BiologyBelum ada peringkat
- The History Behind HDAC Inhibitor Success.20140724.013715Dokumen2 halamanThe History Behind HDAC Inhibitor Success.20140724.013715pvctwine86Belum ada peringkat
- Neurobiology of Circadian Rhythm RegulationDokumen10 halamanNeurobiology of Circadian Rhythm RegulationAgus AgueroBelum ada peringkat
- Molecular Regulations of Circadian Rhythm and Implications For Physiology and DiseasesDokumen20 halamanMolecular Regulations of Circadian Rhythm and Implications For Physiology and Diseasesngthathu.taBelum ada peringkat
- Neuronal Information TransferDari EverandNeuronal Information TransferArthur KarlinBelum ada peringkat
- Neurosciences Research: Volume 4Dari EverandNeurosciences Research: Volume 4S EhrenpreisBelum ada peringkat
- Cold Spring Harb Perspect Biol-2012-Blakely-A005595Dokumen25 halamanCold Spring Harb Perspect Biol-2012-Blakely-A005595Md. Ahsan-Ul BariBelum ada peringkat
- Martin Biel Et Al - Hyperpolarization-Activated Cation Channels: From Genes To FunctionDokumen40 halamanMartin Biel Et Al - Hyperpolarization-Activated Cation Channels: From Genes To FunctionFedrmBelum ada peringkat
- The Neural Circuit of Orexin (Hypocretin) : Maintaining Sleep and WakefulnessDokumen12 halamanThe Neural Circuit of Orexin (Hypocretin) : Maintaining Sleep and WakefulnessvipereejayBelum ada peringkat
- TMP AF9 EDokumen1 halamanTMP AF9 EnithiananthiBelum ada peringkat
- The Circadian System: A Regulatory Feedback Network of Periphery and BrainDokumen12 halamanThe Circadian System: A Regulatory Feedback Network of Periphery and BrainOscarMavilaHerediaBelum ada peringkat
- Hypothalamic Astrocyte Development and Physiology For Neuroprogesterone Induction of The Luteinizing Hormone SurgeDokumen12 halamanHypothalamic Astrocyte Development and Physiology For Neuroprogesterone Induction of The Luteinizing Hormone SurgehfathiardiBelum ada peringkat
- Cushing Syndrome From J Physiol. 2015Dokumen14 halamanCushing Syndrome From J Physiol. 2015no hagoBelum ada peringkat
- 1.kandel Squire NeuroscienceDokumen18 halaman1.kandel Squire NeuroscienceManuel RomeroBelum ada peringkat
- Behavioural Brain Research: Lisa A. Briand, Julie A. BlendyDokumen6 halamanBehavioural Brain Research: Lisa A. Briand, Julie A. BlendyCarlos Eduardo Novoa PalomáBelum ada peringkat
- Mechanism of DyspneaDokumen6 halamanMechanism of Dyspneafenty nisaBelum ada peringkat
- Neurosecretory CellDokumen10 halamanNeurosecretory CellDr Sanjeeb Kumar Dey BaidyaBelum ada peringkat
- Sepsis-Induced Brain Dysfunction: C. Guidoux, T. Sharshar, and D. AnnaneDokumen8 halamanSepsis-Induced Brain Dysfunction: C. Guidoux, T. Sharshar, and D. AnnaneIdris IkhsanBelum ada peringkat
- Biomolecules: Discovery of The First Neurotransmitter Receptor: The Acetylcholine Nicotinic ReceptorDokumen15 halamanBiomolecules: Discovery of The First Neurotransmitter Receptor: The Acetylcholine Nicotinic ReceptorJana CardosoBelum ada peringkat
- Cholinesterases: A Histochemical Contribution to the Solution of Some Functional ProblemsDari EverandCholinesterases: A Histochemical Contribution to the Solution of Some Functional ProblemsBelum ada peringkat
- Cummings 1930Dokumen4 halamanCummings 1930Budiono Gunawan SutantoBelum ada peringkat
- Acute Otitis Media: Case ReportDokumen1 halamanAcute Otitis Media: Case Reportmei risantiBelum ada peringkat
- Daftar PustakaDokumen1 halamanDaftar Pustakamei risantiBelum ada peringkat
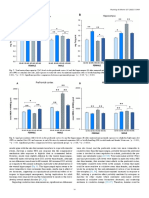
- Diagram Sampel Kortisol Dan Stres Pada TinnitusDokumen8 halamanDiagram Sampel Kortisol Dan Stres Pada Tinnitusmei risantiBelum ada peringkat
- Treatment of Otomycosis Due To Aspergillus Niger With Tolnaftate PDFDokumen3 halamanTreatment of Otomycosis Due To Aspergillus Niger With Tolnaftate PDFMei Risanti SiraitBelum ada peringkat
- Jurnal Reading OtomikosisDokumen4 halamanJurnal Reading OtomikosisTria JuwitaBelum ada peringkat
- Otomycosis ChildrenDokumen3 halamanOtomycosis ChildrenMei Risanti SiraitBelum ada peringkat
- Physiology of Labor PDFDokumen68 halamanPhysiology of Labor PDFAJ MendozaBelum ada peringkat
- James M. Greenblatt, Kelly Brogan-Integrative Therapies For Depression - Redefining Models For Assessment, Treatment and Prevention-CRC Press (2015)Dokumen538 halamanJames M. Greenblatt, Kelly Brogan-Integrative Therapies For Depression - Redefining Models For Assessment, Treatment and Prevention-CRC Press (2015)Werner Heisenberg100% (2)
- Sexual Dimorphism Spatial Learning Metabolism Stress Diet (10-14)Dokumen5 halamanSexual Dimorphism Spatial Learning Metabolism Stress Diet (10-14)Camila Alejandra Inostroza RiveraBelum ada peringkat
- Los Efectos Biológicos Del Trauma InfantilDokumen39 halamanLos Efectos Biológicos Del Trauma InfantilFelipe Cáceres RiquelmeBelum ada peringkat
- Endocrinology Test ReviewDokumen5 halamanEndocrinology Test ReviewHermela GhebremichaelBelum ada peringkat
- Korte 2005Dokumen36 halamanKorte 2005CellaBelum ada peringkat
- Cfa Aromatic Psychodynamics 2018Dokumen40 halamanCfa Aromatic Psychodynamics 2018Agnes_Agnes_AgnesBelum ada peringkat
- Traumatic Stress - Effects On The Brain PDFDokumen17 halamanTraumatic Stress - Effects On The Brain PDFDaniel Londoño GuzmánBelum ada peringkat
- Frontiers in Neuroendocrinology: SciencedirectDokumen15 halamanFrontiers in Neuroendocrinology: SciencedirectFrancisco Javier Luza RamosBelum ada peringkat
- TISSUEDBDokumen8 halamanTISSUEDBArum Purnama SariBelum ada peringkat
- CHROUSOS, George 2009 - Stress and Disorders of The Stress SystemDokumen8 halamanCHROUSOS, George 2009 - Stress and Disorders of The Stress SystemRafael MagalhãesBelum ada peringkat
- EBOOK 978 0123750976 Handbook of Neuroendocrinology Download Full Chapter PDF KindleDokumen61 halamanEBOOK 978 0123750976 Handbook of Neuroendocrinology Download Full Chapter PDF Kindlejuan.speight804100% (34)
- Newport, D.J. and Nemeroff, C.B. Neurobiology of Posttraumatic Stress Disorder. Cognitive Neuroscience JAARTALDokumen8 halamanNewport, D.J. and Nemeroff, C.B. Neurobiology of Posttraumatic Stress Disorder. Cognitive Neuroscience JAARTALg10564433Belum ada peringkat
- Association Between Psychosocial Stress and Hypertension: A Systematic Review and Meta-AnalysisDokumen9 halamanAssociation Between Psychosocial Stress and Hypertension: A Systematic Review and Meta-AnalysisGabriela SiqueiraBelum ada peringkat
- Prenatal Maternal Stress Effects on Pregnancy and Child DevelopmentDokumen12 halamanPrenatal Maternal Stress Effects on Pregnancy and Child Developmentmelpomena81Belum ada peringkat
- The Role of Hormone in ParturitionDokumen36 halamanThe Role of Hormone in ParturitionDiana HayatiBelum ada peringkat
- Crash Course Anatomy and Physiology EndoccrineDokumen9 halamanCrash Course Anatomy and Physiology EndoccrineReem SleemBelum ada peringkat
- Meat ProductionDokumen139 halamanMeat ProductionArooma KhalidBelum ada peringkat
- Post Traumatic Stress Disorder The Neurobiological Impact of Psychological TraumaDokumen17 halamanPost Traumatic Stress Disorder The Neurobiological Impact of Psychological TraumaMariana VilcaBelum ada peringkat
- Pengaruh Senam Maryam Terhadap Tingkat Stres Dan Kadar Kortisol Serum Pada Ibu Hamil PrimiparaDokumen10 halamanPengaruh Senam Maryam Terhadap Tingkat Stres Dan Kadar Kortisol Serum Pada Ibu Hamil PrimiparaTeniadsvnrBelum ada peringkat
- Toward A Neuroscience of AttachmentDokumen34 halamanToward A Neuroscience of AttachmentIoana UngurianuBelum ada peringkat
- Pathophysiology of DepressionDokumen44 halamanPathophysiology of DepressionArmando Marín FloresBelum ada peringkat
- Neurochemistry of MusicDokumen15 halamanNeurochemistry of MusicLisa CerezoBelum ada peringkat
- Stress and Skin Disorders, 2017Dokumen257 halamanStress and Skin Disorders, 2017Elisa Carrera50% (4)
- Back Massage Therapy To Reduce Low Back Pain in PRDokumen4 halamanBack Massage Therapy To Reduce Low Back Pain in PRTian Nopita SariBelum ada peringkat
- Essentials of Abnormal Psychology 7th Edition Durand Test BankDokumen30 halamanEssentials of Abnormal Psychology 7th Edition Durand Test Banknhiamandat4uvr100% (24)
- Crisis Intervention Model for Child Protective ServicesDokumen22 halamanCrisis Intervention Model for Child Protective ServicesKali HartvigsenBelum ada peringkat
- General and Specific Avoidance Coping: The Development and Validation of A New ScaleDokumen253 halamanGeneral and Specific Avoidance Coping: The Development and Validation of A New ScaleChandanaBelum ada peringkat
- Anxiety: Insights Into Signs, Symptoms, Etiology, Pathophysiology, and TreatmentDokumen13 halamanAnxiety: Insights Into Signs, Symptoms, Etiology, Pathophysiology, and TreatmentMichael Judika Deardo PurbaBelum ada peringkat
- The Potential Role of Hypocortisolism in The Pathophysiology of Stress-Related Bodily DisordersDokumen35 halamanThe Potential Role of Hypocortisolism in The Pathophysiology of Stress-Related Bodily DisordersAugusto Felipe Benavides MejiasBelum ada peringkat