Anda mungkin juga menyukai
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (895)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (74)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Dencover ClaimformDokumen4 halamanDencover Claimformd763106Belum ada peringkat
- NP3. Nursing Board Exam November 2008 Answer KeyDokumen12 halamanNP3. Nursing Board Exam November 2008 Answer KeyAlyssa Mier Dacua Patalinjug100% (1)
- Previous View Home: Dimensions Performance SpecificationDokumen124 halamanPrevious View Home: Dimensions Performance SpecificationIntan HaddadBelum ada peringkat
- Phytochemicals: Extraction Methods, Basic Structures and Mode of Action As Potential Chemotherapeutic AgentsDokumen33 halamanPhytochemicals: Extraction Methods, Basic Structures and Mode of Action As Potential Chemotherapeutic AgentsMacDonald OkwudiliChukwu Stain100% (1)
- Article 7 Teacher As Actor: Future English Teachers' Training in The Natural Approach Using DramaDokumen13 halamanArticle 7 Teacher As Actor: Future English Teachers' Training in The Natural Approach Using DramaIntan HaddadBelum ada peringkat
- Lateral Lingual Swellings and One Medial Swelling, The Tuberculum ImparDokumen1 halamanLateral Lingual Swellings and One Medial Swelling, The Tuberculum ImparIntan HaddadBelum ada peringkat
- 04 Graf GoutDokumen20 halaman04 Graf GoutIntan HaddadBelum ada peringkat
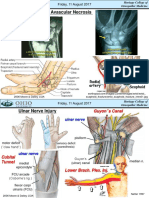
- Scaphoid Fracture & Avascular Necrosis: Friday, 11 August 2017Dokumen4 halamanScaphoid Fracture & Avascular Necrosis: Friday, 11 August 2017Intan HaddadBelum ada peringkat
- Effect of Physiotherapy Rehabilitation On Volkmann Ischemic Contracture-A Case StudyDokumen2 halamanEffect of Physiotherapy Rehabilitation On Volkmann Ischemic Contracture-A Case StudyIntan HaddadBelum ada peringkat
- American Society of Hematology 2018 Guidelines For Management of Venous Thromboembolism Optimal Management of Anticoagulation TherapyDokumen35 halamanAmerican Society of Hematology 2018 Guidelines For Management of Venous Thromboembolism Optimal Management of Anticoagulation TherapyBladimir CentenoBelum ada peringkat
- Gordana Pavliša, Marina Labor, Hrvoje Puretić, Ana Hećimović, Marko Jakopović, Miroslav SamaržijaDokumen12 halamanGordana Pavliša, Marina Labor, Hrvoje Puretić, Ana Hećimović, Marko Jakopović, Miroslav SamaržijaAngelo GarinoBelum ada peringkat
- Influenza Virus Research and Funding Opportunities by John StambasDokumen10 halamanInfluenza Virus Research and Funding Opportunities by John StambasGCEIDBelum ada peringkat
- Toronto Notes Nephrology 2015 10Dokumen1 halamanToronto Notes Nephrology 2015 10JUSASBBelum ada peringkat
- Communicable Disease Nursing Clinical FocusDokumen2 halamanCommunicable Disease Nursing Clinical Focusrceponelas1127Belum ada peringkat
- Effectiveness of Hand Wash and Sanitizer: COVID19: Bulletin of Pure & Applied Sciences-Zoology January 2020Dokumen5 halamanEffectiveness of Hand Wash and Sanitizer: COVID19: Bulletin of Pure & Applied Sciences-Zoology January 2020CarolineBelum ada peringkat
- Iodine DeficiencyDokumen27 halamanIodine DeficiencysunielgowdaBelum ada peringkat
- Quiz RespiratoryDokumen123 halamanQuiz RespiratoryMedShare100% (23)
- 1 Hemoglobinopathies BookDokumen26 halaman1 Hemoglobinopathies BookRhoda Lom-ocBelum ada peringkat
- M1-Medicine MCQs-2004-UNILORINDokumen4 halamanM1-Medicine MCQs-2004-UNILORINIfabiyi OlaniyiBelum ada peringkat
- Funda 1Dokumen8 halamanFunda 1Aijem RyanBelum ada peringkat
- UGIB Prob Sec To BPUD Vs Gastric Mass Alzheimer's DiseaseDokumen7 halamanUGIB Prob Sec To BPUD Vs Gastric Mass Alzheimer's DiseaseMonique Angela Turingan GanganBelum ada peringkat
- Outcome of ERCP in KCGHDokumen6 halamanOutcome of ERCP in KCGHAliA.RamadhanBelum ada peringkat
- Diarrhea in ChildrenDokumen42 halamanDiarrhea in ChildrenIPNATC NEPALBelum ada peringkat
- P Blood Group SystemDokumen2 halamanP Blood Group SystemMiguel SabillenaBelum ada peringkat
- Opalescence Boost 40 PercentDokumen4 halamanOpalescence Boost 40 PercentAnthony GamarraBelum ada peringkat
- A Colour Atlas of FundosDokumen132 halamanA Colour Atlas of FundosMohammed HosenBelum ada peringkat
- Hand, Foot and Mouth Disease What Is It?Dokumen2 halamanHand, Foot and Mouth Disease What Is It?leslie_macasaetBelum ada peringkat
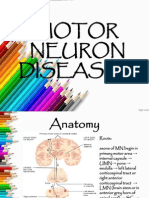
- MNDDokumen28 halamanMNDGayle VillarizaBelum ada peringkat
- Mechanism of HyperventilationDokumen1 halamanMechanism of HyperventilationEdio PathicBelum ada peringkat
- Hospital ListDokumen3 halamanHospital ListShoaib MalikBelum ada peringkat
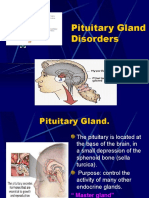
- Pituitary Gland DisordersDokumen80 halamanPituitary Gland DisordersNang KhamBelum ada peringkat
- Drug Classificati ON Mechanism of Action Indications Contraindications Side Effects/ Adverse Effects Nursing ConsiderationsDokumen3 halamanDrug Classificati ON Mechanism of Action Indications Contraindications Side Effects/ Adverse Effects Nursing ConsiderationsFranz Earl Niño AlbesaBelum ada peringkat
- Artificial Hip Joint/Hip Replacement: Muhammad Akmal Qaisar 08-PWMCT-0056 Mechanical Engg DesignDokumen18 halamanArtificial Hip Joint/Hip Replacement: Muhammad Akmal Qaisar 08-PWMCT-0056 Mechanical Engg DesignAkmalBelum ada peringkat
- TabunDokumen24 halamanTabunMiftakul SururiBelum ada peringkat
- Diagnosisandtreatment Approachestoa"Gummy Smile": Harry Dym,, Robert Pierre IiDokumen9 halamanDiagnosisandtreatment Approachestoa"Gummy Smile": Harry Dym,, Robert Pierre Ii刘雨樵Belum ada peringkat
- Pleomorphic AdenomaDokumen4 halamanPleomorphic AdenomaRobins DhakalBelum ada peringkat
- Journal 4Dokumen5 halamanJournal 4nana nurdahliaBelum ada peringkat