Anda mungkin juga menyukai
- Satouh 2018Dokumen11 halamanSatouh 2018Bioinformatica UCSMBelum ada peringkat
- Fertilizacion Revision - EspañolDokumen8 halamanFertilizacion Revision - EspañolAsistente IRP RESTREPOBelum ada peringkat
- Mecanismo Fecundacion HumanaDokumen8 halamanMecanismo Fecundacion HumanaDarwin A Quispe CruzBelum ada peringkat
- Salazar-Cartín J 2019 Biología de La Capacitación EspermáticaDokumen11 halamanSalazar-Cartín J 2019 Biología de La Capacitación EspermáticaJorge Alberto Salazar CartinBelum ada peringkat
- Desarrollo Regulativo y MosaicoDokumen14 halamanDesarrollo Regulativo y MosaicoEvelynBelum ada peringkat
- Capacitación espermática: cambios bioquímicos y fisiológicos requeridos para la fecundaciónDokumen11 halamanCapacitación espermática: cambios bioquímicos y fisiológicos requeridos para la fecundaciónRoger Machacuay ZavalaBelum ada peringkat
- Embriologia Parte 2Dokumen65 halamanEmbriologia Parte 2Valerie Mendoza MendezBelum ada peringkat
- GametogénesisDokumen4 halamanGametogénesisAndrea Mesa Arrieta100% (1)
- La Reproducción en AnfibiosDokumen2 halamanLa Reproducción en AnfibiosEvelynBelum ada peringkat
- Fisiologia Capacitacion Espermatica 1 PDFDokumen7 halamanFisiologia Capacitacion Espermatica 1 PDFRichard BedónBelum ada peringkat
- Evaluación Mitocondrial en GametosDokumen13 halamanEvaluación Mitocondrial en GametosAlesu RodríguezBelum ada peringkat
- Unidad 1 EmbriologiaDokumen99 halamanUnidad 1 EmbriologiaanaelisagarciaBelum ada peringkat
- Embrion Aborto Pildora Del Dia SiguienteDokumen62 halamanEmbrion Aborto Pildora Del Dia SiguientePercy Zapata Mendo.Belum ada peringkat
- Sinteis ConceptualesDokumen348 halamanSinteis ConceptualesBRAYAN BARRAZA MACIASBelum ada peringkat
- Trabajo de Embriologia Semana 1Dokumen6 halamanTrabajo de Embriologia Semana 1Lupita lazo carmenBelum ada peringkat
- Rsa 03109Dokumen5 halamanRsa 03109Luis ArteagaBelum ada peringkat
- Informe de Anahi 222Dokumen13 halamanInforme de Anahi 222katherinegb2006Belum ada peringkat
- Factores que influyen en la capacitación espermáticaDokumen3 halamanFactores que influyen en la capacitación espermáticaRenzo Cortez PachecoBelum ada peringkat
- Resumen Artículo 6 JLCRDokumen3 halamanResumen Artículo 6 JLCRGeorge Luis Colunga RamirezBelum ada peringkat
- Generalidades de Cestodos - Recursos en Parasitología - Unam (Checked)Dokumen4 halamanGeneralidades de Cestodos - Recursos en Parasitología - Unam (Checked)pixoguiasBelum ada peringkat
- Práctica 9Dokumen6 halamanPráctica 9serafinpris2Belum ada peringkat
- The Key Role of Vitamin A in Spermatogenesis (ESPAÑOL)Dokumen8 halamanThe Key Role of Vitamin A in Spermatogenesis (ESPAÑOL)oliverdelgado323Belum ada peringkat
- Fisiologia Inicio Espontáneo de Trabajo de Parto PDFDokumen13 halamanFisiologia Inicio Espontáneo de Trabajo de Parto PDFAlfredoBelum ada peringkat
- Erizo de Mar LaboratorioDokumen9 halamanErizo de Mar LaboratorioValery Sue Barreda OrtizBelum ada peringkat
- Anatomia y Funcion Del Nicho de Celulas MadreDokumen14 halamanAnatomia y Funcion Del Nicho de Celulas MadreRosmary HornaBelum ada peringkat
- Cap 04Dokumen9 halamanCap 04joaBelum ada peringkat
- Fol I Culo GenesisDokumen9 halamanFol I Culo GenesisAnayatzy LaraBelum ada peringkat
- Apunte 3 - Gestacion y PlacentacionDokumen22 halamanApunte 3 - Gestacion y Placentacioncrvallejos100% (3)
- Articulo de Biología - En.esDokumen10 halamanArticulo de Biología - En.esCamilo Andres Paternina BedoyaBelum ada peringkat
- Practica 5 Desarrolo Morfologivco HumanoDokumen4 halamanPractica 5 Desarrolo Morfologivco HumanoLiz Giorgina Araoz TuanamaBelum ada peringkat
- Taller 2Dokumen5 halamanTaller 2Fernando Garcia MoralesBelum ada peringkat
- Sintesis Conceptuales Del Capitulo 1Dokumen28 halamanSintesis Conceptuales Del Capitulo 1Luz CristinaBelum ada peringkat
- p53 y su papel en el epitelio superficial del ovarioDokumen33 halamanp53 y su papel en el epitelio superficial del ovarioNestor lorenzoBelum ada peringkat
- Consulta 6Dokumen6 halamanConsulta 6Angy PavónBelum ada peringkat
- FECUNDACIÓNDokumen7 halamanFECUNDACIÓNjulissagonzalezzuritaBelum ada peringkat
- Topofosis-control-cambiosDokumen2 halamanTopofosis-control-cambiosErick Jean Melgar Gutierrez0% (1)
- Toxoplasma Gondii - Biologia MolecularDokumen3 halamanToxoplasma Gondii - Biologia MolecularAlex Valle SotoBelum ada peringkat
- Capacitacion EspermaticaDokumen11 halamanCapacitacion EspermaticaSharon MosquedaBelum ada peringkat
- Editorial 2Dokumen2 halamanEditorial 2EVELIN JOHANA LEMA HEREDIABelum ada peringkat
- FecundaciónDokumen7 halamanFecundaciónElizabeth Mondragon hoyosBelum ada peringkat
- Zona PelúcidaDokumen6 halamanZona PelúcidaCamila PlataBelum ada peringkat
- Histologia Grupo 2Dokumen15 halamanHistologia Grupo 2Mirely BustamanteBelum ada peringkat
- Cuestionario EmbriologíaDokumen89 halamanCuestionario EmbriologíaDavenia SaldañaBelum ada peringkat
- Infertilidad MasculinaDokumen23 halamanInfertilidad MasculinaLuis Eduardo Soto GuardoBelum ada peringkat
- Preguntas 6 7 8Dokumen11 halamanPreguntas 6 7 8Yordan Smith BustosBelum ada peringkat
- Avances en El Conocimiento de La Espermatogénesis HumanaDokumen14 halamanAvances en El Conocimiento de La Espermatogénesis HumanaJenniffer RonquilloBelum ada peringkat
- Guía de Fecundación Humana.Dokumen9 halamanGuía de Fecundación Humana.CAMILA CONSTANZA ROJAS OLMOSBelum ada peringkat
- 02 Pregenesis y FecundacionDokumen14 halaman02 Pregenesis y FecundacionIrene C. CruzBelum ada peringkat
- Amanda KR Nunes (2013) Análisis Morfológica e Funcional Del Proceso Espermatogénico en CobayosDokumen13 halamanAmanda KR Nunes (2013) Análisis Morfológica e Funcional Del Proceso Espermatogénico en CobayosDavid Choque HuamanBelum ada peringkat
- Tutoria de ObstetriciaDokumen33 halamanTutoria de ObstetriciaJoshenka Garay IntriagoBelum ada peringkat
- 4TA A 8VA SEMANA A PDFDokumen12 halaman4TA A 8VA SEMANA A PDFMaryori Treyci ILasaca ValenzuelaBelum ada peringkat
- Control Genetico Del Desarrollo EmbrionarioDokumen3 halamanControl Genetico Del Desarrollo EmbrionarioJonathan VzlaBelum ada peringkat
- The Cytoskeleton in Spermatogenesis (ESPAÑOL)Dokumen20 halamanThe Cytoskeleton in Spermatogenesis (ESPAÑOL)oliverdelgado323Belum ada peringkat
- Laboratorio 1Dokumen9 halamanLaboratorio 1moisesBelum ada peringkat
- III Clase ObstetriciaDokumen51 halamanIII Clase ObstetriciaRaúl Alexis Menudo ProblemaBelum ada peringkat
- Copia de Morfología de Gónadas - Observación y Caracterización - de La Ossa, Ospino, Trespalacios-1Dokumen7 halamanCopia de Morfología de Gónadas - Observación y Caracterización - de La Ossa, Ospino, Trespalacios-1sdelaossaBelum ada peringkat
- Tumores de Celulas GigantesDokumen13 halamanTumores de Celulas GigantesAna Lucía RománBelum ada peringkat
- Fusión CelularDokumen7 halamanFusión CelularLABORATORIO HOSPITAL SAN RICARDO PAMPURIBelum ada peringkat
- Inmunolocalización de Sonic Hedgehog en el desarrollo embrio-fetal de ratonesDokumen6 halamanInmunolocalización de Sonic Hedgehog en el desarrollo embrio-fetal de ratonesFlor Elena ToxquiBelum ada peringkat
- Anexo 1Dokumen2 halamanAnexo 1Brandon Chahua ObregonBelum ada peringkat
- ECOLOGIADokumen2 halamanECOLOGIABrandon Chahua Obregon0% (1)
- Bienvenido PDFDokumen5 halamanBienvenido PDFRaquel Pérez PérezBelum ada peringkat
- Liderazgo en Medicina - MedigraphicDokumen9 halamanLiderazgo en Medicina - MedigraphicBrandon Chahua ObregonBelum ada peringkat
- Juan SociodramaDokumen1 halamanJuan SociodramaBrandon Chahua ObregonBelum ada peringkat
- CRONOGRAMADokumen1 halamanCRONOGRAMABrandon Chahua ObregonBelum ada peringkat
- Anexo 1Dokumen2 halamanAnexo 1Brandon Chahua ObregonBelum ada peringkat
- Resumen XDDokumen5 halamanResumen XDBrandon Chahua ObregonBelum ada peringkat
- PRÁCTICADokumen1 halamanPRÁCTICABrandon Chahua ObregonBelum ada peringkat
- 01 SegmentosDokumen3 halaman01 SegmentosBrandon Chahua ObregonBelum ada peringkat
- Dialnet CalidadYLiderazgoEnMedicina 4701391 PDFDokumen11 halamanDialnet CalidadYLiderazgoEnMedicina 4701391 PDFJuarez 3710Belum ada peringkat
- Liderazgo en Medicina - MedigraphicDokumen9 halamanLiderazgo en Medicina - MedigraphicBrandon Chahua ObregonBelum ada peringkat
- PLANTILLA-DE-EXPOSICION BrandonDokumen5 halamanPLANTILLA-DE-EXPOSICION BrandonBrandon Chahua ObregonBelum ada peringkat
- Resumen 1Dokumen4 halamanResumen 1Brandon Chahua ObregonBelum ada peringkat
- Anquilostiomosis 2019Dokumen12 halamanAnquilostiomosis 2019Brandon Chahua ObregonBelum ada peringkat
- Cy A 4to Primara La CélulaDokumen1 halamanCy A 4to Primara La CélulaBrandon Chahua ObregonBelum ada peringkat
- Malla Curricular Medicina Humana Plan 04Dokumen4 halamanMalla Curricular Medicina Humana Plan 04CANAL DE ING. ELECTRONICA UNJFSCBelum ada peringkat
- PLANTILLA-DE-EXPOSICION BrandonDokumen5 halamanPLANTILLA-DE-EXPOSICION BrandonBrandon Chahua ObregonBelum ada peringkat
- 4tl4s Its MedicinafullDokumen92 halaman4tl4s Its MedicinafullCézhar Navarro Pardavé100% (1)
- Cuestionario 111111Dokumen6 halamanCuestionario 111111Brandon Chahua ObregonBelum ada peringkat
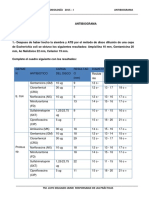
- Cuestionario #05 AntibiogramaDokumen5 halamanCuestionario #05 AntibiogramaBrandon Chahua ObregonBelum ada peringkat
- Espectrofotometría: Funcionamiento y aplicaciones claveDokumen2 halamanEspectrofotometría: Funcionamiento y aplicaciones claveBrandon Chahua Obregon100% (1)
- Qué Es Una EnzimaDokumen30 halamanQué Es Una EnzimaBrandon Chahua ObregonBelum ada peringkat
- Resultados 1Dokumen1 halamanResultados 1Brandon Chahua ObregonBelum ada peringkat
- INDICEDokumen2 halamanINDICEBrandon Chahua ObregonBelum ada peringkat
- Cuestionario N 9Dokumen5 halamanCuestionario N 9Brandon Chahua ObregonBelum ada peringkat
- Cuestionario #07Dokumen2 halamanCuestionario #07Brandon Chahua ObregonBelum ada peringkat
- Qué Es Una EnzimaDokumen6 halamanQué Es Una EnzimaBrandon Chahua Obregon100% (1)
- Monografía de La LepraDokumen23 halamanMonografía de La Leprasamcr000100% (4)
- Diagnostico Inicial Panaderia La ComisanaDokumen36 halamanDiagnostico Inicial Panaderia La ComisanaDjOscarGamixBelum ada peringkat
- Repaso EcorregionesDokumen3 halamanRepaso EcorregionesMarco JimenezBelum ada peringkat
- Comidas Tipicas de ChihuahuaDokumen6 halamanComidas Tipicas de ChihuahuaLuuiz LoveeBelum ada peringkat
- DemonDokumen86 halamanDemonMisael SalazarBelum ada peringkat
- Sindrome de Craneo HipertensivoDokumen11 halamanSindrome de Craneo HipertensivoDalia GómezBelum ada peringkat
- CIENCIAS DE LA NATURALEZA Tema 1Dokumen1 halamanCIENCIAS DE LA NATURALEZA Tema 1Belén Valdivia CifuentesBelum ada peringkat
- Contactos Gays Escort Pollon en MadridDokumen3 halamanContactos Gays Escort Pollon en MadridEroticoBelum ada peringkat
- Fonetica y OratoriaDokumen16 halamanFonetica y OratoriaTania Amparo Bolados MarambioBelum ada peringkat
- Aulonocara SPDokumen2 halamanAulonocara SPBorja González-CaminoBelum ada peringkat
- Guia Médica - Envenenamiento Por Animales PonzoñososDokumen15 halamanGuia Médica - Envenenamiento Por Animales PonzoñososadsommxBelum ada peringkat
- Paolicchi, Cerdá y Kameniecki. Reflejos Del Recién NacidoDokumen5 halamanPaolicchi, Cerdá y Kameniecki. Reflejos Del Recién NacidopatrikaleBelum ada peringkat
- Tarea 2 U1 Barrera Barraza Victor AntonioDokumen3 halamanTarea 2 U1 Barrera Barraza Victor AntonioVictor Barrera BarrazaBelum ada peringkat
- Balleneros Guipuzcoanos en Las Matanzas de Ballenas de Galicia y Asturias - Siglos Xvi y XviiDokumen35 halamanBalleneros Guipuzcoanos en Las Matanzas de Ballenas de Galicia y Asturias - Siglos Xvi y XviiPSVBelum ada peringkat
- Tutoria Fichas 2° Grado PrimariaDokumen38 halamanTutoria Fichas 2° Grado PrimariaBRUNO DíAZ "BATMAN Bca"100% (3)
- RECUERDO PERDIDO: LA ESCULTURA DE AMESDokumen3 halamanRECUERDO PERDIDO: LA ESCULTURA DE AMESjulio cesar silvaBelum ada peringkat
- Fisiologia de Renal Parte 1Dokumen26 halamanFisiologia de Renal Parte 1MariPaty LorenzoBelum ada peringkat
- Osopolarpedia ComDokumen5 halamanOsopolarpedia ComAriana PagliaBelum ada peringkat
- 1plan de Charla de Paternidad Matenriada ResponsableDokumen9 halaman1plan de Charla de Paternidad Matenriada ResponsableMadely Choroco Salvatierra100% (1)
- Mecanismo de Trabajo de PartoDokumen42 halamanMecanismo de Trabajo de PartoMariafernadaZea100% (2)
- Cuidados nutricionales para colostomíaDokumen1 halamanCuidados nutricionales para colostomíaAna Torrejon100% (1)
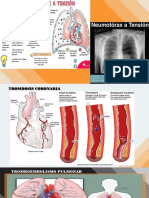
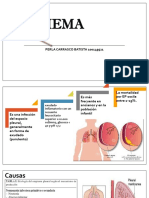
- Empiema Mi Expo Grupo1Dokumen21 halamanEmpiema Mi Expo Grupo1Perla Carrasco BatistaBelum ada peringkat
- LenguajeDokumen176 halamanLenguajeKrlos Sani100% (1)
- Analisis Cefalometrico de Kim (Meaw)Dokumen14 halamanAnalisis Cefalometrico de Kim (Meaw)David Sanchez Martinez100% (8)
- Grupo 3 Navarrete Correa Barreto CrispinDokumen38 halamanGrupo 3 Navarrete Correa Barreto Crispinjoseph nazarioBelum ada peringkat
- 03 - Cirugía Del Pie Equinovaro CongénitoDokumen11 halaman03 - Cirugía Del Pie Equinovaro CongénitoLuis Angel Ponce Torres100% (1)
- Articles-82309 Recurso PDFDokumen2 halamanArticles-82309 Recurso PDFignacio salvador diaz donosoBelum ada peringkat
- CUESTIONARIO PARA VALORAR CONOCIMIENTOS - OdtDokumen5 halamanCUESTIONARIO PARA VALORAR CONOCIMIENTOS - Odtorlaned2Belum ada peringkat
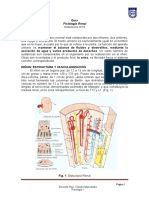
- Guía Fisiología Renal Enfermería 2019Dokumen17 halamanGuía Fisiología Renal Enfermería 2019Borreguinho OrregoBelum ada peringkat
- Vias BiliaresDokumen4 halamanVias BiliarespyzamuBelum ada peringkat