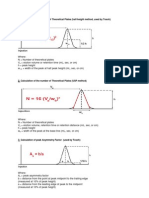
Anda mungkin juga menyukai
- 001 7-Steps-Template-of-Financial-Statements-and-Modeling-and-ValuationDokumen113 halaman001 7-Steps-Template-of-Financial-Statements-and-Modeling-and-ValuationAbhinav VermaBelum ada peringkat
- Ex Vitro Rooting and Ex Vitro Acclimation by Using The 'Float Hydroponics' Technique - AlexfiraonthewebDokumen10 halamanEx Vitro Rooting and Ex Vitro Acclimation by Using The 'Float Hydroponics' Technique - AlexfiraonthewebBrij Mohan SinghBelum ada peringkat
- Introduction Part For Onion Root Tip KaryotypingDokumen2 halamanIntroduction Part For Onion Root Tip Karyotypingciaralim04100% (2)
- Rental Home Financial Projection TemplateDokumen61 halamanRental Home Financial Projection TemplateMinh NguyenBelum ada peringkat
- California Tenan'Ts-Landlord-Tenant Rights and ResponsibilitiesDokumen124 halamanCalifornia Tenan'Ts-Landlord-Tenant Rights and ResponsibilitiesfuziontvBelum ada peringkat
- Basic Financial Package Template (With COGS)Dokumen47 halamanBasic Financial Package Template (With COGS)Rukundo PatrickBelum ada peringkat
- GMP CannabisDokumen49 halamanGMP CannabisAna Julia Sotomayor Adrada100% (1)
- Crop Profitability CalculatorDokumen12 halamanCrop Profitability CalculatorAnupam BaliBelum ada peringkat
- Standards Science Behind The EarthboxDokumen3 halamanStandards Science Behind The Earthboxapi-253362431Belum ada peringkat
- Extending Shelf Life of Guava Fruits by Mint Oil and UVC TreatmentsDokumen9 halamanExtending Shelf Life of Guava Fruits by Mint Oil and UVC TreatmentsIJEAB JournalBelum ada peringkat
- How To Avoid To Lose Your Shirt in AquacultureDokumen4 halamanHow To Avoid To Lose Your Shirt in AquacultureRoberto MarchesiniBelum ada peringkat
- Aeroponic Farming: The Future of AgricultureDokumen3 halamanAeroponic Farming: The Future of AgricultureiiNirvanaii DIYBelum ada peringkat
- Godrej Meadows BrochureDokumen42 halamanGodrej Meadows BrochurePrashant Joshi100% (1)
- Financial Projections TemplateDokumen214 halamanFinancial Projections TemplateReselyn PalabinoBelum ada peringkat
- Section 14 - Pesticides Management in Jamaica PDFDokumen63 halamanSection 14 - Pesticides Management in Jamaica PDFNatalieBelum ada peringkat
- 8th WAC Brochure - 09072018Dokumen13 halaman8th WAC Brochure - 09072018Shubham Ballav SemwalBelum ada peringkat
- An Integrated Fish Culture and Vegetable Hydroponics Production SystemDokumen8 halamanAn Integrated Fish Culture and Vegetable Hydroponics Production Systemkriskee13Belum ada peringkat
- One Acre Aquaponics Farm by ENochDokumen29 halamanOne Acre Aquaponics Farm by ENochEnoch Aaron R'GasanaBelum ada peringkat
- Closing Nutrient Cycles in Cities Urban Vermiponics (PDFDrive)Dokumen24 halamanClosing Nutrient Cycles in Cities Urban Vermiponics (PDFDrive)aallffaa2009Belum ada peringkat
- Annexure 2 - Sample Financial Model TemplateDokumen135 halamanAnnexure 2 - Sample Financial Model TemplateGurvinder SinghBelum ada peringkat
- Aquaculture in GhanaDokumen6 halamanAquaculture in GhanaInternational Aquafeed magazineBelum ada peringkat
- Poultry Farming - 4 LITEDokumen18 halamanPoultry Farming - 4 LITEAnonymous tW1zTL2ltBelum ada peringkat
- List of Tools - Mushroom CultivationDokumen1 halamanList of Tools - Mushroom CultivationVillamor NiezBelum ada peringkat
- 5 Year Financial Plan Calculation ForsendingDokumen33 halaman5 Year Financial Plan Calculation ForsendingBernard James TandangBelum ada peringkat
- The Cost of Producing Potted Orchids Hawaiian Agricultural ProductsDokumen4 halamanThe Cost of Producing Potted Orchids Hawaiian Agricultural ProductsStefana LnBelum ada peringkat
- NYC Study of Waste-to-Energy OptionsDokumen209 halamanNYC Study of Waste-to-Energy OptionsCity Limits (New York)Belum ada peringkat
- Effect of Firm Ripe Plantain Fruit Flour Addition On The Chemical, Sensory and Microbial Quality of Fura PowderDokumen7 halamanEffect of Firm Ripe Plantain Fruit Flour Addition On The Chemical, Sensory and Microbial Quality of Fura PowderAnonymous fKrx2rFBelum ada peringkat
- 33 of The Healthiest Foods On Earth Si 520Dokumen60 halaman33 of The Healthiest Foods On Earth Si 520FeiFei WangBelum ada peringkat
- IC Detailed Financial Projections Template 8821 UpdatedDokumen27 halamanIC Detailed Financial Projections Template 8821 UpdatedRozh SammedBelum ada peringkat
- Microbiological Safety Testing of CannabisDokumen54 halamanMicrobiological Safety Testing of CannabisGdanskJam JamBelum ada peringkat
- Royalty AccountingDokumen10 halamanRoyalty AccountingsmlingwaBelum ada peringkat
- Budget For Greenhouse TomatoesDokumen8 halamanBudget For Greenhouse Tomatoessina981Belum ada peringkat
- Column Efficiency TestingDokumen3 halamanColumn Efficiency TestingvamsiBelum ada peringkat
- Swiggy - 5 Year To Go SustainableDokumen1 halamanSwiggy - 5 Year To Go SustainableKirana RetailBelum ada peringkat
- PPM Cannamimetics White PaperDokumen33 halamanPPM Cannamimetics White PaperChadBerginBelum ada peringkat
- Abatin Wellness Center 2Dokumen77 halamanAbatin Wellness Center 2New Jersey marijuana documents100% (1)
- Extraction Methods, Chemical Characterizations and Biological Activities ofDokumen9 halamanExtraction Methods, Chemical Characterizations and Biological Activities ofJUAN C. OVIEDO LOPERABelum ada peringkat
- Yadala, Soumya - Design of Raceway Ponds For Biomass ProductionDokumen21 halamanYadala, Soumya - Design of Raceway Ponds For Biomass ProductionJorge Antonio Arce OrtizBelum ada peringkat
- How To Garlic Genger PasteDokumen23 halamanHow To Garlic Genger PasteAmmad Ahmed KhanBelum ada peringkat
- SprocesDokumen4 halamanSprocesFernando SanchezBelum ada peringkat
- Plant Tissue Culture An Alternative For Production of Useful MetabolitesDokumen79 halamanPlant Tissue Culture An Alternative For Production of Useful MetabolitesNaiem Mubarak100% (1)
- Water Use in Cannabis AgricultureDokumen2 halamanWater Use in Cannabis AgricultureAdam ForgieBelum ada peringkat
- Hemp Business ProposalDokumen7 halamanHemp Business ProposalAnthony Ulloa0% (1)
- Bill Snow Financial Model 2004-03-09Dokumen35 halamanBill Snow Financial Model 2004-03-09nsadnanBelum ada peringkat
- Avocado Fruit Quality Issues On The International MarketDokumen14 halamanAvocado Fruit Quality Issues On The International MarketViviana Delgado100% (1)
- 7 Steps Template of Financial Statements and Modeling and ValuationDokumen108 halaman7 Steps Template of Financial Statements and Modeling and ValuationErica Lenn VillagraciaBelum ada peringkat
- Balance Sheet TemplateDokumen4 halamanBalance Sheet TemplateSolomon Ahimbisibwe100% (1)
- Reported Species of Mushroom in Pakistan From Upto Year 1997Dokumen19 halamanReported Species of Mushroom in Pakistan From Upto Year 1997AlijanBelum ada peringkat
- Rathi, V., Herbal Wine A ReviewDokumen7 halamanRathi, V., Herbal Wine A ReviewBrian Mukti NugrohoBelum ada peringkat
- PLANT TISSUE CULTURE (Sinan-188801230)Dokumen11 halamanPLANT TISSUE CULTURE (Sinan-188801230)LOTUSBelum ada peringkat
- Material and MethodsDokumen26 halamanMaterial and MethodsAkhilaBelum ada peringkat
- Biotechnology BIOT3113 TC Lab Manual 2012Dokumen15 halamanBiotechnology BIOT3113 TC Lab Manual 2012Sakile' GarlandBelum ada peringkat
- Blue Green AlgaeDokumen39 halamanBlue Green AlgaeAnuj RajBelum ada peringkat
- Research Advisor: Hod:: DR - Vimala Beera, Asst - Professor DR - Ch.V.V.Satyanarayana, Professor & HeadDokumen49 halamanResearch Advisor: Hod:: DR - Vimala Beera, Asst - Professor DR - Ch.V.V.Satyanarayana, Professor & HeadBharat GuduruBelum ada peringkat
- Biochem Laboratory 2Dokumen2 halamanBiochem Laboratory 2Lorainne Angel U. MolinaBelum ada peringkat
- Cell Culturing EXP 4 Plant Tissue Culture PDFDokumen8 halamanCell Culturing EXP 4 Plant Tissue Culture PDFPhú NguyễnBelum ada peringkat
- 06 - Chapter 3Dokumen35 halaman06 - Chapter 3VijayakumarBelum ada peringkat
- Title: Preparation of Medium and Growth Hormone Stock Solutions. Objectives: 1) To Learn On How To Prepare Concentrated Stocks SolutionsDokumen8 halamanTitle: Preparation of Medium and Growth Hormone Stock Solutions. Objectives: 1) To Learn On How To Prepare Concentrated Stocks SolutionsCharles Raj100% (1)
- SCP Production of Single Cell Protein From Agrowaste Using Rhodococcus OpacusDokumen19 halamanSCP Production of Single Cell Protein From Agrowaste Using Rhodococcus OpacusTran Le Thanh NgocBelum ada peringkat
- The Power of WordsDokumen10 halamanThe Power of Wordsciaralim04Belum ada peringkat
- Gods Dream by Bo SanchezDokumen28 halamanGods Dream by Bo SanchezPauline Joanne Arceo100% (1)
- A Starfish Can Wipe Out ReefsDokumen20 halamanA Starfish Can Wipe Out Reefsciaralim04100% (2)
- Discussion On The Results For KaryotypingDokumen1 halamanDiscussion On The Results For Karyotypingciaralim04Belum ada peringkat
- Philosophy of BiologyDokumen47 halamanPhilosophy of BiologyDanny De Leon100% (2)
- Health - Diabetes Research PaperDokumen7 halamanHealth - Diabetes Research Paperapi-299727107Belum ada peringkat
- Virology SummaryDokumen3 halamanVirology SummaryvictorBelum ada peringkat
- Introduction To Medical Microbiology 2023Dokumen93 halamanIntroduction To Medical Microbiology 2023Samuel fikaduBelum ada peringkat
- Thesis Complete PDFDokumen269 halamanThesis Complete PDFshjahsjanshaBelum ada peringkat
- (Methods in Molecular Biology 823) Stacy M. Cowherd (Auth.), Virginia Espina, Lance A. Liotta (Eds.) - Molecular Profiling - Methods and Protocols (2012, Humana Press)Dokumen462 halaman(Methods in Molecular Biology 823) Stacy M. Cowherd (Auth.), Virginia Espina, Lance A. Liotta (Eds.) - Molecular Profiling - Methods and Protocols (2012, Humana Press)ashutosh pandey100% (1)
- Hb-2560-003 HB Qiaamp PWRFCL Pro Dna Kit 0220 WWDokumen24 halamanHb-2560-003 HB Qiaamp PWRFCL Pro Dna Kit 0220 WWMysaraSamyǮesawyBelum ada peringkat
- Chapter 30 Homework Packet KeyDokumen14 halamanChapter 30 Homework Packet Keyrosaroncero0% (4)
- D&A: 28082020 NIH Grant NoticeDokumen386 halamanD&A: 28082020 NIH Grant NoticeD&A Investigations, Inc.Belum ada peringkat
- Contoh Review Artikel (Presenter)Dokumen2 halamanContoh Review Artikel (Presenter)Tri SuwandiBelum ada peringkat
- Theories of Developmental PsychologyDokumen10 halamanTheories of Developmental PsychologyAbenezer YohannesBelum ada peringkat
- Evaluation of Meropenem, Imipenem and Ertapenem Impregnated MacConkey Agar Plates For The Detection of Carbapenem Resistant EnterobacteriaceaeDokumen8 halamanEvaluation of Meropenem, Imipenem and Ertapenem Impregnated MacConkey Agar Plates For The Detection of Carbapenem Resistant EnterobacteriaceaeInternational Medical PublisherBelum ada peringkat
- Full Download Test Bank For Biology of Humans Concepts Applications and Issues 5th Edition Goodenough PDF Full ChapterDokumen36 halamanFull Download Test Bank For Biology of Humans Concepts Applications and Issues 5th Edition Goodenough PDF Full Chapterplagate.fanega.872zlw100% (15)
- Importances of Reproduction OrganismsDokumen17 halamanImportances of Reproduction OrganismsPORCHE CAYENNE MACAN GTSBelum ada peringkat
- Quarter 1 - GenBio 1-Week 1 (23-24)Dokumen41 halamanQuarter 1 - GenBio 1-Week 1 (23-24)Adrienne Skyle SibalaBelum ada peringkat
- ENS ReportDokumen13 halamanENS ReportAirastormBelum ada peringkat
- PiXL Knowledge Test QUESTIONS - AQA B1 CORE Science - Legacy (2016 and 2017)Dokumen6 halamanPiXL Knowledge Test QUESTIONS - AQA B1 CORE Science - Legacy (2016 and 2017)Mrs S BakerBelum ada peringkat
- Bacteria From PrescottDokumen19 halamanBacteria From PrescottYekitaSBelum ada peringkat
- 6.8 Exchange of Gases in The LungsDokumen22 halaman6.8 Exchange of Gases in The LungsyousafBelum ada peringkat
- Scooby's Suggested Solutions For The 2014 VCE Biology Exam - Short AnswerDokumen6 halamanScooby's Suggested Solutions For The 2014 VCE Biology Exam - Short AnswerRite10210Belum ada peringkat
- General Biology 00 Burl U of TDokumen612 halamanGeneral Biology 00 Burl U of TAndreiClaudiuBelum ada peringkat
- Technical Guidelines Pinus HeldreichiiDokumen6 halamanTechnical Guidelines Pinus HeldreichiiBakir GanićBelum ada peringkat
- TellmeGen Interpretation ManualDokumen22 halamanTellmeGen Interpretation Manualhello goodbyeBelum ada peringkat
- Mendelian GeneticsDokumen6 halamanMendelian GeneticsMaria Danica100% (2)
- Cornell Notes CH 7Dokumen2 halamanCornell Notes CH 7Aaron WongBelum ada peringkat
- Extended Validity of Reserve Lists 2020Dokumen11 halamanExtended Validity of Reserve Lists 2020yiotaliapiBelum ada peringkat
- TN - Gram Positive Vs Gram Negative - Article - JL - V2ksDokumen5 halamanTN - Gram Positive Vs Gram Negative - Article - JL - V2ksNGÂN NGUYỄN HOÀNG TRÚCBelum ada peringkat
- Unit 1:: Learning OutcomesDokumen15 halamanUnit 1:: Learning OutcomesJhenny Lyn CamanoBelum ada peringkat
- Plant Cell:: Ultra Structure and Functions of Typical Plant and Animal CellDokumen8 halamanPlant Cell:: Ultra Structure and Functions of Typical Plant and Animal CellMaleehaBelum ada peringkat
- Plant Reproductive StructuresDokumen59 halamanPlant Reproductive StructuresJames Daniel OmalinBelum ada peringkat
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessDari EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessBelum ada peringkat
- Why We Die: The New Science of Aging and the Quest for ImmortalityDari EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityPenilaian: 4 dari 5 bintang4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDari EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDari EverandTales from Both Sides of the Brain: A Life in NeurosciencePenilaian: 3 dari 5 bintang3/5 (18)
- Gut: the new and revised Sunday Times bestsellerDari EverandGut: the new and revised Sunday Times bestsellerPenilaian: 4 dari 5 bintang4/5 (392)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDari EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RacePenilaian: 4.5 dari 5 bintang4.5/5 (516)
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincDari EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincPenilaian: 3.5 dari 5 bintang3.5/5 (137)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Dari EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Penilaian: 4 dari 5 bintang4/5 (378)
- World of Wonders: In Praise of Fireflies, Whale Sharks, and Other AstonishmentsDari EverandWorld of Wonders: In Praise of Fireflies, Whale Sharks, and Other AstonishmentsPenilaian: 4 dari 5 bintang4/5 (223)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDari EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeBelum ada peringkat
- Dark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniverseDari EverandDark Matter and the Dinosaurs: The Astounding Interconnectedness of the UniversePenilaian: 3.5 dari 5 bintang3.5/5 (69)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDari EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedPenilaian: 4 dari 5 bintang4/5 (11)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDari EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouPenilaian: 4.5 dari 5 bintang4.5/5 (62)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDari EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsPenilaian: 4 dari 5 bintang4/5 (5)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperDari EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperPenilaian: 4.5 dari 5 bintang4.5/5 (15)
- Who's in Charge?: Free Will and the Science of the BrainDari EverandWho's in Charge?: Free Will and the Science of the BrainPenilaian: 4 dari 5 bintang4/5 (65)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDari EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionPenilaian: 4 dari 5 bintang4/5 (811)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDari Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessPenilaian: 4 dari 5 bintang4/5 (33)
- Water: The Epic Struggle for Wealth, Power, and CivilizationDari EverandWater: The Epic Struggle for Wealth, Power, and CivilizationPenilaian: 3.5 dari 5 bintang3.5/5 (37)
- The Other End of the Leash: Why We Do What We Do Around DogsDari EverandThe Other End of the Leash: Why We Do What We Do Around DogsPenilaian: 5 dari 5 bintang5/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDari EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorBelum ada peringkat
- Good Without God: What a Billion Nonreligious People Do BelieveDari EverandGood Without God: What a Billion Nonreligious People Do BelievePenilaian: 4 dari 5 bintang4/5 (66)