Anda mungkin juga menyukai
- The Toxicity of Caffein: An experimental study on different species of animalsDari EverandThe Toxicity of Caffein: An experimental study on different species of animalsBelum ada peringkat
- 395 08 11 ArticleDokumen8 halaman395 08 11 ArticleRidha Surya NugrahaBelum ada peringkat
- Jcinvest00248 0177 PDFDokumen11 halamanJcinvest00248 0177 PDFluis morales garzaBelum ada peringkat
- Phosphorylase Activity of Skeletal Muscle Extracts 1955 Krebs e FisherDokumen8 halamanPhosphorylase Activity of Skeletal Muscle Extracts 1955 Krebs e Fisherdavi2002ramalhoBelum ada peringkat
- J. Biol. Chem.-1918-Harding-575-80Dokumen7 halamanJ. Biol. Chem.-1918-Harding-575-80Vincenzo Paolo FraddosioBelum ada peringkat
- Effects of Insulin-Like Growth Factor On Nitrogen Balance During Hypoxic ExposureDokumen7 halamanEffects of Insulin-Like Growth Factor On Nitrogen Balance During Hypoxic ExposurelaurentiaBelum ada peringkat
- AttachmentDokumen4 halamanAttachmentLawliet FardiBelum ada peringkat
- 1 s2.0 S0022227520390441 MainDokumen4 halaman1 s2.0 S0022227520390441 MainVanessa StepaniaBelum ada peringkat
- 1962 Cholesterol Metabolism in GerbilDokumen5 halaman1962 Cholesterol Metabolism in GerbilVitoria LimaBelum ada peringkat
- Action of Dinitrophenol and Insulin in Accelerating The Metab of Ethyl Alcohol 1935Dokumen4 halamanAction of Dinitrophenol and Insulin in Accelerating The Metab of Ethyl Alcohol 1935Zaharia MariusBelum ada peringkat
- Tryptophan Appetite Healthy: DepletionDokumen6 halamanTryptophan Appetite Healthy: DepletionRifqi Hamdani PasaribuBelum ada peringkat
- Investigation of Irisin's Role in Pubertal Onset Physiology in Female RatsDokumen6 halamanInvestigation of Irisin's Role in Pubertal Onset Physiology in Female RatsonderhuseyinbasBelum ada peringkat
- Jejum IntermitenteDokumen11 halamanJejum IntermitenteJohn Talmid De Yeshua HamashiachBelum ada peringkat
- Karatsoreos Et Al 2010Dokumen11 halamanKaratsoreos Et Al 2010zacharymweil100% (1)
- Hildegarde Esther Allen: City of Medical AND Division OF OFDokumen7 halamanHildegarde Esther Allen: City of Medical AND Division OF OFJulien Patrick CebrianBelum ada peringkat
- Antidiabetic and Long-Term Effects of Elaeocarpus GrandiflorusDokumen12 halamanAntidiabetic and Long-Term Effects of Elaeocarpus GrandiflorusAgus FakhrudinBelum ada peringkat
- Of Caffeine, Theophylline and Theobromine On Scheduled Controlled Responding in RatsDokumen4 halamanOf Caffeine, Theophylline and Theobromine On Scheduled Controlled Responding in RatsMukilan RanjithBelum ada peringkat
- Islet Cell Aislamiento en Experimental D, L-Etionina Pancreatitis en Perro1-S2.0-0022480485900381-MainDokumen9 halamanIslet Cell Aislamiento en Experimental D, L-Etionina Pancreatitis en Perro1-S2.0-0022480485900381-MainCarlos VargasBelum ada peringkat
- Research ArticleDokumen9 halamanResearch Articlehemanth kumarBelum ada peringkat
- In Vitro Study of Acetylcholine and Histamine Induced Contractions in Colon and Rectum of Adult and Neonate RatsDokumen10 halamanIn Vitro Study of Acetylcholine and Histamine Induced Contractions in Colon and Rectum of Adult and Neonate RatsErum JanBelum ada peringkat
- 2003 Butler Lucy Insulin GH - IGF1Dairy (3320)Dokumen13 halaman2003 Butler Lucy Insulin GH - IGF1Dairy (3320)Sueños De LunaBelum ada peringkat
- F?e Ing R Gimes de CR Bed O. 5 5 I : Nature. Vol. 215. SeptemberDokumen2 halamanF?e Ing R Gimes de CR Bed O. 5 5 I : Nature. Vol. 215. SeptemberhanoomleeBelum ada peringkat
- ArtifDokumen15 halamanArtifIstván PortörőBelum ada peringkat
- Efecto de CetonaDokumen9 halamanEfecto de CetonaGiusi CaforioBelum ada peringkat
- 9048 FullDokumen5 halaman9048 FullFelia AlyciaBelum ada peringkat
- Fact #1: Wellness & Fitness-Body OptimizationDokumen13 halamanFact #1: Wellness & Fitness-Body OptimizationCheeken CharliBelum ada peringkat
- Effect of Intermittent Fasting and Refeeding On Insulin Action in Healthy MenDokumen9 halamanEffect of Intermittent Fasting and Refeeding On Insulin Action in Healthy Mendgh3Belum ada peringkat
- Effects of Clenbuterol and Propranolol On Muscle Mass Evidence That Clenbuterol Stimulates Muscle Adrenoceptors To Induce HypertrophyDokumen7 halamanEffects of Clenbuterol and Propranolol On Muscle Mass Evidence That Clenbuterol Stimulates Muscle Adrenoceptors To Induce HypertrophyDANNIELI GALVEZBelum ada peringkat
- Insuficiencia Pancreática Exocrina Canina Tratada Con Extracto Pancreático PorcinoDokumen4 halamanInsuficiencia Pancreática Exocrina Canina Tratada Con Extracto Pancreático PorcinoAndreaBelum ada peringkat
- Nutritional and Protein: Albumin Metabolism: Effect of The State The IntakeDokumen15 halamanNutritional and Protein: Albumin Metabolism: Effect of The State The IntakeDio PattersonBelum ada peringkat
- Study of Antidiabetic Activity of Hiptage Benghalensis (L) KurzDokumen8 halamanStudy of Antidiabetic Activity of Hiptage Benghalensis (L) KurzIOSR Journal of PharmacyBelum ada peringkat
- Gutman Et Al 2011cDokumen4 halamanGutman Et Al 2011cRoee GutmanBelum ada peringkat
- Effects of Boldenone Undecylenate oDokumen7 halamanEffects of Boldenone Undecylenate ogrzewo666Belum ada peringkat
- Studies On The Metabolism of Semen. 5. Citric Acid in SemenDokumen9 halamanStudies On The Metabolism of Semen. 5. Citric Acid in Semenrohit.jaruplaBelum ada peringkat
- Non-Toxicity of IV Injected Perfluorocarbon Oxygen Carrier in An Animal Model of Liver Regeneration Following Surgical InjuryDokumen9 halamanNon-Toxicity of IV Injected Perfluorocarbon Oxygen Carrier in An Animal Model of Liver Regeneration Following Surgical InjuryIstván PortörőBelum ada peringkat
- Role Protein Degradation: of in The Growth of Livers After A Nutritional ShiftDokumen6 halamanRole Protein Degradation: of in The Growth of Livers After A Nutritional ShiftEnrique Pardo PerezBelum ada peringkat
- Effects of Pair-Feeding and Growth Hormone Treatment On Obese Transgenic RatsDokumen5 halamanEffects of Pair-Feeding and Growth Hormone Treatment On Obese Transgenic RatsThanh LeBelum ada peringkat
- Research Article: The Extract of Herbal Medicines Activates AMP-Activated Protein Kinase in Diet-Induced Obese RatsDokumen9 halamanResearch Article: The Extract of Herbal Medicines Activates AMP-Activated Protein Kinase in Diet-Induced Obese RatsJoshBelum ada peringkat
- Intermittent Fasting and Refeeding On Insulin Dependant MenDokumen10 halamanIntermittent Fasting and Refeeding On Insulin Dependant MenTyrone PenningBelum ada peringkat
- Original Article Evidence of Anti-Obesity Effects of The Pomegranate Leaf Extract in High-Fat Diet Induced Obese MiceDokumen7 halamanOriginal Article Evidence of Anti-Obesity Effects of The Pomegranate Leaf Extract in High-Fat Diet Induced Obese MiceAisy Savira anizarBelum ada peringkat
- Philippe Ducrotté and Pierre Déchelotte Moïse Coëffier, Sophie Claeyssens, Bernadette Hecketsweiler, Alain LavoinneDokumen9 halamanPhilippe Ducrotté and Pierre Déchelotte Moïse Coëffier, Sophie Claeyssens, Bernadette Hecketsweiler, Alain LavoinneMoonsferatuBelum ada peringkat
- Fuels, Hormones, and Liver Metabolism Term and During The Early Postnatal Period in The RatDokumen11 halamanFuels, Hormones, and Liver Metabolism Term and During The Early Postnatal Period in The RatAnna IzabelBelum ada peringkat
- Fasting REEDokumen5 halamanFasting REEAimone FerriBelum ada peringkat
- Journal of Dental Research: Influence of Gastric Acidity On Fluoride Absorption in RatsDokumen5 halamanJournal of Dental Research: Influence of Gastric Acidity On Fluoride Absorption in RatsRia AlfadinaBelum ada peringkat
- Intermittent Fasting Induces Hypothalamic Modifications Resulting in Low Feeding Efficiency, Low Body Mass and Overating 2014Dokumen11 halamanIntermittent Fasting Induces Hypothalamic Modifications Resulting in Low Feeding Efficiency, Low Body Mass and Overating 2014Camila VidigalBelum ada peringkat
- Tissue Modifications in Monkeys As Related To Absorption, Distribution, and Excretion of Polychlorinated BiphenylsDokumen10 halamanTissue Modifications in Monkeys As Related To Absorption, Distribution, and Excretion of Polychlorinated BiphenylsSh1vaBelum ada peringkat
- Cruciferous Vegetables and Cancer PreventionDokumen22 halamanCruciferous Vegetables and Cancer Preventionpranayzenith5485Belum ada peringkat
- Clinical: Influence of Pharmacokinetics Bioavailability Highly Purified Beef DependentDokumen5 halamanClinical: Influence of Pharmacokinetics Bioavailability Highly Purified Beef Dependentsstrumello7395Belum ada peringkat
- Kapeghian Et Al 1983Dokumen3 halamanKapeghian Et Al 1983John MaddenBelum ada peringkat
- Vandenbraak2013 - CoagulaciónDokumen7 halamanVandenbraak2013 - CoagulaciónLeti PeraltaBelum ada peringkat
- Spontaneous 24-h Ghrelin Secretion Pattern in FastDokumen7 halamanSpontaneous 24-h Ghrelin Secretion Pattern in Fastmv.Belum ada peringkat
- The Plasma Sugar, Free Fatty Acid, Cortisol, and Growth Hormone Response Insulin. I. in Control SubjectsDokumen8 halamanThe Plasma Sugar, Free Fatty Acid, Cortisol, and Growth Hormone Response Insulin. I. in Control SubjectsYng TangBelum ada peringkat
- Spesialis Penyakit Dalam 2 (Additive Effects of Rebamipide Plus Proton Pump PDFDokumen5 halamanSpesialis Penyakit Dalam 2 (Additive Effects of Rebamipide Plus Proton Pump PDFfildzah amalyaBelum ada peringkat
- Che Cura 2010Dokumen8 halamanChe Cura 2010camila andrea rivera diazBelum ada peringkat
- Ginger PublishedDokumen8 halamanGinger PublishedFatimoh AbdulsalamBelum ada peringkat
- Hepatic Effects of Halothane, Iso Urane or Sevo Urane Anaesthesia in DogsDokumen4 halamanHepatic Effects of Halothane, Iso Urane or Sevo Urane Anaesthesia in DogstoblerumBelum ada peringkat
- Anti-Diabetic Activities of Fucosterol From: Pelvetia SiliquosaDokumen3 halamanAnti-Diabetic Activities of Fucosterol From: Pelvetia SiliquosaLeandreux OcasionBelum ada peringkat
- Tannis As AstringentDokumen5 halamanTannis As Astringentmueen hashmiBelum ada peringkat
- The Ghrelin Receptor: in Vivo Characterization of High Basal Signaling FromDokumen11 halamanThe Ghrelin Receptor: in Vivo Characterization of High Basal Signaling FromFernando Menéndez AliagaBelum ada peringkat
- Cyprinus Carpio Fed Diets With DifferentDokumen6 halamanCyprinus Carpio Fed Diets With DifferentjosekinBelum ada peringkat
- 12 Desalination Advance Water Treatment Economics FinancingDokumen169 halaman12 Desalination Advance Water Treatment Economics FinancingFlavia ZanetteBelum ada peringkat
- Pi Molecular Orbital DiagramDokumen9 halamanPi Molecular Orbital DiagramIon DebianBelum ada peringkat
- Furanic Compounds PDFDokumen6 halamanFuranic Compounds PDFAbdullah GhannamBelum ada peringkat
- 1251 Weighing On An Analytical BalanceDokumen6 halaman1251 Weighing On An Analytical Balancenonick18Belum ada peringkat
- Princípio de Babinet 1Dokumen3 halamanPrincípio de Babinet 1josh16kalebBelum ada peringkat
- Chapter Sixteen Cell SignalingDokumen96 halamanChapter Sixteen Cell SignalingRu LiliBelum ada peringkat
- PseudoephedrineDokumen2 halamanPseudoephedrineSiti Marwasofa100% (3)
- Emerson.2012. The Microbial Ferrous Wheel: Iron Cycling in Terrestrial, Freshwater, and Marine Environments - BookDokumen217 halamanEmerson.2012. The Microbial Ferrous Wheel: Iron Cycling in Terrestrial, Freshwater, and Marine Environments - BookMilena NovaBelum ada peringkat
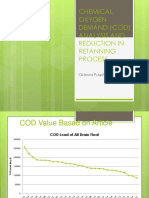
- COD Anaysis and ReductionDokumen13 halamanCOD Anaysis and ReductionelaBelum ada peringkat
- Pressurised Deaerator Head-Technical Information PDFDokumen2 halamanPressurised Deaerator Head-Technical Information PDFel doctolBelum ada peringkat
- Environmental Aspects PDFDokumen31 halamanEnvironmental Aspects PDFShyam SinghBelum ada peringkat
- Lecture3 Example ShellandtuberatingcalculationsDokumen4 halamanLecture3 Example Shellandtuberatingcalculationsunknown8787Belum ada peringkat
- Concrete Reinforcement and Glass Fibre Reinforced PolymerDokumen9 halamanConcrete Reinforcement and Glass Fibre Reinforced PolymerchanakaBelum ada peringkat
- BRITT - Hi-Bay LN6101 BrochureDokumen1 halamanBRITT - Hi-Bay LN6101 BrochureRenato Abalos100% (1)
- Biofouling Potential and Environmental Factors of Seawater ADokumen20 halamanBiofouling Potential and Environmental Factors of Seawater ATrieLanyPutriYuliananingrumBelum ada peringkat
- SG1001 ChemlokAdhesivesDokumen6 halamanSG1001 ChemlokAdhesivesAnonymous e0kwBYSaBelum ada peringkat
- Tips On ExamDokumen12 halamanTips On ExamB ruikarBelum ada peringkat
- Backwards Heat Equation InfoDokumen1 halamanBackwards Heat Equation InfoMelissa Marie HernandezBelum ada peringkat
- App Guide - Xl16iapg01enb TraneDokumen20 halamanApp Guide - Xl16iapg01enb TraneAngel Quique MeanaBelum ada peringkat
- Colloids and Surfaces A: Physicochemical and Engineering AspectsDokumen9 halamanColloids and Surfaces A: Physicochemical and Engineering AspectsChí ChungBelum ada peringkat
- Tool CodeDokumen16 halamanTool CodeAhmed BouBelum ada peringkat
- PSV 430066 PDFDokumen41 halamanPSV 430066 PDFErikikoBelum ada peringkat
- 2 Effect of Heat On SaltDokumen3 halaman2 Effect of Heat On SaltJacelynBelum ada peringkat
- Dimitri: Ivanovich MendeleevDokumen13 halamanDimitri: Ivanovich MendeleevJilu Mary AntonyBelum ada peringkat
- Vanadio NMRDokumen7 halamanVanadio NMRsergioodin4851Belum ada peringkat
- 8.4 Performance Qualification Protocol For Dispensing BoothDokumen13 halaman8.4 Performance Qualification Protocol For Dispensing BoothArej Ibrahim AbulailBelum ada peringkat
- GT3Dokumen14 halamanGT3VenkateshPrasadBelum ada peringkat
- ElectrochemistryDokumen24 halamanElectrochemistryZainul AbedeenBelum ada peringkat
- FT-IR Spectrophotometric Analysis of Acetylsalicylic Acid and Its Pharmaceutical Formulations.Dokumen7 halamanFT-IR Spectrophotometric Analysis of Acetylsalicylic Acid and Its Pharmaceutical Formulations.luna_mirabelleBelum ada peringkat
- Nagendra G M: Teaching ExperienceDokumen3 halamanNagendra G M: Teaching Experiencenagendra_gmBelum ada peringkat