Anda mungkin juga menyukai
- I Congreso Colombiano de Filosofía- Estética, fenomenología y hermenéutica - Volumen IDari EverandI Congreso Colombiano de Filosofía- Estética, fenomenología y hermenéutica - Volumen IBelum ada peringkat
- La Colonia de Hormigas Como Organismo (1911) - William Morton WheelerDokumen25 halamanLa Colonia de Hormigas Como Organismo (1911) - William Morton WheeleranaelfoBelum ada peringkat
- Constelaciones para niños: ¡La forma más divertida de aprender sobre las estrellas, descubrir la magia del sistema solar y observar los astros como un astrónomo!Dari EverandConstelaciones para niños: ¡La forma más divertida de aprender sobre las estrellas, descubrir la magia del sistema solar y observar los astros como un astrónomo!Belum ada peringkat
- La Carta Del Preste JuanDokumen38 halamanLa Carta Del Preste JuanSara Daniela Garcia DiazBelum ada peringkat
- Atlan Con Razon o Sin EllaDokumen4 halamanAtlan Con Razon o Sin ElladenisenajBelum ada peringkat
- Entre dos mundos: la antropología radical de Paul StollerDari EverandEntre dos mundos: la antropología radical de Paul StollerBelum ada peringkat
- Freud, Sigmund - Los Actos Obsesivos y Las Practicas ReligiosasDokumen7 halamanFreud, Sigmund - Los Actos Obsesivos y Las Practicas ReligiosasHernando HerreroBelum ada peringkat
- El silencio de Dios: Relato de una supervivienteDari EverandEl silencio de Dios: Relato de una supervivienteBelum ada peringkat
- Art Pranayama Bks IyengarDokumen3 halamanArt Pranayama Bks Iyengarpato_aballayBelum ada peringkat
- OntoPsiquis - Más allá del eneagrama y el psicoanálisis: OntoPsiquis - Más allá del eneagrama y el psicoanálisis, #1Dari EverandOntoPsiquis - Más allá del eneagrama y el psicoanálisis: OntoPsiquis - Más allá del eneagrama y el psicoanálisis, #1Belum ada peringkat
- Enrique Eskenazi: Vigencia de La AstrologíaDokumen3 halamanEnrique Eskenazi: Vigencia de La AstrologíaJordiPuigBelum ada peringkat
- Alberto Vital - La Cama de Procusto-Poesía en MovimientoDokumen8 halamanAlberto Vital - La Cama de Procusto-Poesía en MovimientoDalia Fernanda XN TrejoBelum ada peringkat
- Las Puertas de La India - Olivia CattedraDokumen20 halamanLas Puertas de La India - Olivia CattedraJorge Raúl OlguínBelum ada peringkat
- Desarrollo Del Principio Femenino - Sukie ColegraveDokumen9 halamanDesarrollo Del Principio Femenino - Sukie ColegraveCarlos AguilarBelum ada peringkat
- LACAN - Otros Trabajos - de La Psicosis ParanoicaDokumen217 halamanLACAN - Otros Trabajos - de La Psicosis ParanoicaRenato FarabeufBelum ada peringkat
- El Sexo, La Sangre, La MuerteDokumen6 halamanEl Sexo, La Sangre, La MuerteInocencio Verdugo RojoBelum ada peringkat
- Alan Watts Amor y ConocimientoDokumen9 halamanAlan Watts Amor y ConocimientoJosé Pulido Cruz PsicólogoBelum ada peringkat
- Heraclito y Parmenides Maestros de SabiduriaDokumen10 halamanHeraclito y Parmenides Maestros de SabiduriaTiobobyBelum ada peringkat
- La Amada Nocturna de San Juan de La Cruz Se Pudo Haber Llamado Layla Luce Lopez-BaraltDokumen11 halamanLa Amada Nocturna de San Juan de La Cruz Se Pudo Haber Llamado Layla Luce Lopez-BaraltAngelManriqueBelum ada peringkat
- Gaia - Un Hogar para La Psicología Ambiental PDFDokumen34 halamanGaia - Un Hogar para La Psicología Ambiental PDFRolando Alonzo Gutiérrez100% (1)
- Apolo y Los Oscuros Lugares Del SaberDokumen9 halamanApolo y Los Oscuros Lugares Del SaberFebe PaganBelum ada peringkat
- René Guénon - Cartas A Vasile Lovinescu (III)Dokumen11 halamanRené Guénon - Cartas A Vasile Lovinescu (III)caiorossiBelum ada peringkat
- Frithjof Schuon - Miradas Sobre Los Mundos Antiguos PDFDokumen5 halamanFrithjof Schuon - Miradas Sobre Los Mundos Antiguos PDFquinbusflestrinBelum ada peringkat
- Elogio de La Dificultad - Estanislao ZuletaDokumen3 halamanElogio de La Dificultad - Estanislao ZuletaPaulo César MesaBelum ada peringkat
- El Mito Cuenta Una Historia SagradaDokumen2 halamanEl Mito Cuenta Una Historia SagradaIara Ortiz100% (1)
- Cristianismo y Astrología MONTERODokumen10 halamanCristianismo y Astrología MONTERORosalía Hernández GarcíaBelum ada peringkat
- Raitzin Eso ExoterismoDokumen5 halamanRaitzin Eso ExoterismoSAÚL PÉREZBelum ada peringkat
- La SerpienteDokumen47 halamanLa Serpientemlph1979Belum ada peringkat
- Teilhard de Chardin Pierre - El Fenomeno HumanoDokumen151 halamanTeilhard de Chardin Pierre - El Fenomeno HumanoMaria Jose Guerrero Arzave100% (1)
- ORIGEN POLAR DE LA TRADICIÓN VÉDICA - Tilak PDFDokumen18 halamanORIGEN POLAR DE LA TRADICIÓN VÉDICA - Tilak PDFEmbosKadoBelum ada peringkat
- Jung y GuénonDokumen4 halamanJung y GuénonJack ShepardBelum ada peringkat
- El Retorno de Las Madres - JJ BachofenDokumen20 halamanEl Retorno de Las Madres - JJ BachofenNube Kusamalut LazzoBelum ada peringkat
- YasnaDokumen3 halamanYasnadon_joe108Belum ada peringkat
- El Alma MasculinaDokumen8 halamanEl Alma MasculinaludussacerBelum ada peringkat
- Emma Jung Marie LouiseDokumen4 halamanEmma Jung Marie LouiseAmarú VanegasBelum ada peringkat
- El Gnosticismo Aún ContemporáneoDokumen6 halamanEl Gnosticismo Aún ContemporáneoSugeyht Jonas Trevelyan-GreyBelum ada peringkat
- Alvarez Lopez, José - El Enigma de Las PiramidesDokumen159 halamanAlvarez Lopez, José - El Enigma de Las PiramidesSantiago ScalercioBelum ada peringkat
- Los Gigantes de VoronezhDokumen4 halamanLos Gigantes de Voronezhmadagascar10Belum ada peringkat
- Burckhardt Titus - Clave Espiritual de La Astrologia MusulmanaDokumen30 halamanBurckhardt Titus - Clave Espiritual de La Astrologia MusulmanaLili Buratti100% (1)
- La Sexualidad, Sacramento EsencialDokumen22 halamanLa Sexualidad, Sacramento EsencialSantiago Christos H D HBelum ada peringkat
- Caliz y La EspadaDokumen20 halamanCaliz y La EspadaSotilllo Yerbes UbencioBelum ada peringkat
- Rene Guenon y Carlos MarxDokumen13 halamanRene Guenon y Carlos MarxGocho Versolari100% (1)
- Anna Kingsford y Edward Maitland - La Via Esoterica de CristoDokumen4 halamanAnna Kingsford y Edward Maitland - La Via Esoterica de Cristoperezru_20006444Belum ada peringkat
- Arsgravis - Arte y Simbolismo - Universidad de Barcelona - LIBROSDokumen14 halamanArsgravis - Arte y Simbolismo - Universidad de Barcelona - LIBROSRintrah ChimallBelum ada peringkat
- Arte y Espiritualidad Ignaciana Liberados para Crear PDFDokumen17 halamanArte y Espiritualidad Ignaciana Liberados para Crear PDFgustavoBelum ada peringkat
- AvestaDokumen3 halamanAvestadon_joe108Belum ada peringkat
- Eros y Logos. Mito y PsiqueDokumen8 halamanEros y Logos. Mito y PsiqueVictor Duque OtalvaroBelum ada peringkat
- Registro de Sueños. Atisbos A La Conciencia Onírica Desde Las Ciencias, Las Artes y La Filosofía PDFDokumen3 halamanRegistro de Sueños. Atisbos A La Conciencia Onírica Desde Las Ciencias, Las Artes y La Filosofía PDFSer NoexistirBelum ada peringkat
- Psicología Analítica de Jung. El Proceso de Individuación.Dokumen3 halamanPsicología Analítica de Jung. El Proceso de Individuación.rapanui62Belum ada peringkat
- La Sabiduría Esotérica de PlatónDokumen4 halamanLa Sabiduría Esotérica de Platónjohnnathan gorsiraBelum ada peringkat
- Tesis Carril19 PDFDokumen288 halamanTesis Carril19 PDFSantiago Posada V.Belum ada peringkat
- Lives in The Shadow (001 140) TaducidoDokumen106 halamanLives in The Shadow (001 140) TaducidoFranco PiacentiniBelum ada peringkat
- El Paso de La Gran RazaDokumen87 halamanEl Paso de La Gran RazaIván Palacios EspañaBelum ada peringkat
- El Gran Juego: Revelación-RevoluciónDokumen14 halamanEl Gran Juego: Revelación-RevoluciónPabloRUizSaavedra100% (1)
- Cultura para La PazDokumen24 halamanCultura para La PazOscar GonzalezBelum ada peringkat
- Alan Watts - La Suprema IdentidadDokumen4 halamanAlan Watts - La Suprema IdentidadXelo Besó MontanerBelum ada peringkat
- J RIES El Sentido de Lo Sagrado en Las Culturas y en Las Religiones Ed Azul Barcelona 2008Dokumen64 halamanJ RIES El Sentido de Lo Sagrado en Las Culturas y en Las Religiones Ed Azul Barcelona 2008Edgar GarcíaBelum ada peringkat
- 58-Bien (El) y El Mal - Las Dos Manos de DiosDokumen1 halaman58-Bien (El) y El Mal - Las Dos Manos de DiosJessica Martinez100% (1)
- Elabora Un Cuadro en El Que Enlistes 5 Características de Los Organismos Unicelulares y 5 de Los PluricelularesDokumen3 halamanElabora Un Cuadro en El Que Enlistes 5 Características de Los Organismos Unicelulares y 5 de Los PluricelularesCrisBelum ada peringkat
- El Sistema de Seguridad de Planta de GasDokumen14 halamanEl Sistema de Seguridad de Planta de GasJaime Garcia Mena100% (1)
- Fusion de Protoplastos Final CompletoDokumen26 halamanFusion de Protoplastos Final CompletoLuis Fernando Ruiz Dulanto100% (1)
- Manual de Laboratorio Botánica 2017-01-30Dokumen108 halamanManual de Laboratorio Botánica 2017-01-30Ana Maria Marin TamayoBelum ada peringkat
- Anon - Manual de Capacitacion en Agricultura Organica PDFDokumen362 halamanAnon - Manual de Capacitacion en Agricultura Organica PDFGabo Castellanos100% (1)
- Practica 3Dokumen13 halamanPractica 3Francisco José Ojeda SanchezBelum ada peringkat
- Capítulo Microfósiles PDFDokumen47 halamanCapítulo Microfósiles PDFSebas EspínBelum ada peringkat
- Diferencia Entre Cactus y SuculentasDokumen4 halamanDiferencia Entre Cactus y SuculentasPatriciasBelum ada peringkat
- Preguntas Generadoras 1Dokumen7 halamanPreguntas Generadoras 1AURA CRISTINA ALMARIO ORTIZBelum ada peringkat
- Trabajo Colaborativo Paso 2 Grupo 25 FinalDokumen15 halamanTrabajo Colaborativo Paso 2 Grupo 25 FinalEdwin López Gutierrez100% (5)
- Triptico Fotosintesis BLANCO Y NEGRODokumen2 halamanTriptico Fotosintesis BLANCO Y NEGROTío Pedro Ríos100% (2)
- Practica 1 Fpo Clasificacion de FrutosDokumen11 halamanPractica 1 Fpo Clasificacion de FrutosBetty Acero ChoqueBelum ada peringkat
- Actividades La Célula IiDokumen3 halamanActividades La Célula Iirestaurante llajaBelum ada peringkat
- Temas 01 Eval. 1 Al 7 BioquímicaDokumen172 halamanTemas 01 Eval. 1 Al 7 BioquímicaJusta CAÑIBANO PALMEROBelum ada peringkat
- Guía de Cultivador PDFDokumen28 halamanGuía de Cultivador PDFreimaxBelum ada peringkat
- Tinción y Reconocimiento de Células Animales, Vegetales y Organismos Celulares - Dana ManuDokumen3 halamanTinción y Reconocimiento de Células Animales, Vegetales y Organismos Celulares - Dana ManuMelissa Arango FlóresBelum ada peringkat
- Cuadro Comparativo Teoría Homóloga y de Interpolación Sobre La Evolución de Las Plantas ContinentalesDokumen2 halamanCuadro Comparativo Teoría Homóloga y de Interpolación Sobre La Evolución de Las Plantas ContinentalesAlonso YanezBelum ada peringkat
- ComunidadDokumen7 halamanComunidadHector Hugo Andrade QuispeBelum ada peringkat
- AA2-U4 Tejidos VegetalesDokumen8 halamanAA2-U4 Tejidos VegetalesEvelyn RomeroBelum ada peringkat
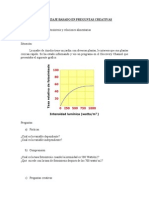
- Preguntas Creativas FotosintesisDokumen4 halamanPreguntas Creativas FotosintesisEli SanhuezaBelum ada peringkat
- Interacciones Planta-Herbicida PAPADokumen24 halamanInteracciones Planta-Herbicida PAPAguidoBelum ada peringkat
- Plantas Medicinales de FerDokumen20 halamanPlantas Medicinales de FerbrakysBelum ada peringkat
- Estrategias de Ciencias Unidad 8 de Octavo GradoDokumen21 halamanEstrategias de Ciencias Unidad 8 de Octavo GradoSamuel Aramis Aramis Mancia Trinidad100% (1)
- Alberto MAssaroDokumen26 halamanAlberto MAssaroFlavio Lagos GutierrezBelum ada peringkat
- Actividad EcologiaDokumen7 halamanActividad EcologiaErik Reyes NVBelum ada peringkat
- Formato Informe de LaboratorioDokumen5 halamanFormato Informe de LaboratorioLuzii OsoriioBelum ada peringkat
- HISTOLOGIADokumen91 halamanHISTOLOGIAGeovannaBelum ada peringkat
- Aminoacidos en Las PlantasDokumen9 halamanAminoacidos en Las PlantasLeslie Caroline Quirohuayo MendozaBelum ada peringkat
- Conceptos de BotánicaDokumen47 halamanConceptos de BotánicaFELIPE RODRIGUEZBelum ada peringkat
- Métodos de Transformación GenéticaDokumen3 halamanMétodos de Transformación Genéticamoend4vBelum ada peringkat
- Unidad 3 Exposicion Equipo 2Dokumen19 halamanUnidad 3 Exposicion Equipo 2Ignacio Pablo LukasBelum ada peringkat