Anda mungkin juga menyukai
- Asimilación Metabólica de Los Alcoholes en Drosophzla: Nutrición YDokumen17 halamanAsimilación Metabólica de Los Alcoholes en Drosophzla: Nutrición YEstefani BlancasBelum ada peringkat
- Metabolism and Interactions: The Chemistry and Biology of Compounds Containing Amino SugarsDari EverandMetabolism and Interactions: The Chemistry and Biology of Compounds Containing Amino SugarsEndre A. BalazsBelum ada peringkat
- Kuznetsova 2006Dokumen14 halamanKuznetsova 2006Fede ZannierBelum ada peringkat
- IAA Protein Conjugadotes 2008Dokumen6 halamanIAA Protein Conjugadotes 2008shady182Belum ada peringkat
- Reproduction in Domestic AnimalsDari EverandReproduction in Domestic AnimalsH. H. ColeBelum ada peringkat
- Symposium On Developmental Genetics: International Congress of GeneticsDokumen10 halamanSymposium On Developmental Genetics: International Congress of GeneticsJohn VargheseBelum ada peringkat
- Distribution and Biological Role: The Chemistry and Biology of Compounds Containing Amino SugarsDari EverandDistribution and Biological Role: The Chemistry and Biology of Compounds Containing Amino SugarsEndre A. BalazsBelum ada peringkat
- Pathogen-Related Yeast (PRY) Proteins and Members of The CAP Superfamily Are Secreted Sterol-Binding Proteins. - 2012 - Choudhary, SchneDokumen6 halamanPathogen-Related Yeast (PRY) Proteins and Members of The CAP Superfamily Are Secreted Sterol-Binding Proteins. - 2012 - Choudhary, SchnemasurBelum ada peringkat
- Abass Toxicology 2011Dokumen10 halamanAbass Toxicology 2011vnq6gd6qbxBelum ada peringkat
- Sintese Fenilhidrazina 1 PDFDokumen12 halamanSintese Fenilhidrazina 1 PDFanon_860281856Belum ada peringkat
- Regulation of Macromolecular Synthesis By Low Molecular Weight MediatorsDari EverandRegulation of Macromolecular Synthesis By Low Molecular Weight MediatorsGebhard KochBelum ada peringkat
- Mechanisms of Saccharide Polymerization and DepolymerizationDari EverandMechanisms of Saccharide Polymerization and DepolymerizationBelum ada peringkat
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967Dari EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterBelum ada peringkat
- Jasmonates Meet Fatty Acids: Functional Analysis of A New Acyl-Coenzyme A Synthetase Family From Arabidopsis ThalianaDokumen17 halamanJasmonates Meet Fatty Acids: Functional Analysis of A New Acyl-Coenzyme A Synthetase Family From Arabidopsis ThalianaFrontiersBelum ada peringkat
- The Shikimate Pathway: Biosynthesis of Natural Products SeriesDari EverandThe Shikimate Pathway: Biosynthesis of Natural Products SeriesBelum ada peringkat
- ODH Review Servier Manuscript 22-831.R1 RevisedDokumen32 halamanODH Review Servier Manuscript 22-831.R1 RevisedTelek AndrsBelum ada peringkat
- FEMS Yeast Research 2019Dokumen8 halamanFEMS Yeast Research 2019carlosniregaladoBelum ada peringkat
- 1.the Role of Lipid Droplets in Metabolic Disease in Rodents and HumansDokumen9 halaman1.the Role of Lipid Droplets in Metabolic Disease in Rodents and HumansShalika AbeysingheBelum ada peringkat
- Biosynthesis of Ergot AlkaloidsDokumen70 halamanBiosynthesis of Ergot AlkaloidspBelum ada peringkat
- Phylogenomic Analysis of Shikimate Dehydrogenases Reveals Functional DiversificationDokumen12 halamanPhylogenomic Analysis of Shikimate Dehydrogenases Reveals Functional DiversificationBreixo HarguindeyBelum ada peringkat
- Physiology of Elasmobranch Fishes: Internal ProcessesDari EverandPhysiology of Elasmobranch Fishes: Internal ProcessesBelum ada peringkat
- Tethered Yohimbine Analogs As Selective Human - Adrenergic Receptor LigandsDokumen10 halamanTethered Yohimbine Analogs As Selective Human - Adrenergic Receptor LigandsMano RatyiBelum ada peringkat
- PAKs, RAC/CDC42 (p21)-activated Kinases: Towards the Cure of Cancer and Other PAK-dependent DiseasesDari EverandPAKs, RAC/CDC42 (p21)-activated Kinases: Towards the Cure of Cancer and Other PAK-dependent DiseasesHiroshi MarutaBelum ada peringkat
- tmpF9AC TMPDokumen7 halamantmpF9AC TMPFrontiersBelum ada peringkat
- Amaranth Proteins As A Source of AntioxiDokumen8 halamanAmaranth Proteins As A Source of AntioxiJherson Gustavo SánchezBelum ada peringkat
- Bacterial lipolytic enzymes classification and propertiesDokumen7 halamanBacterial lipolytic enzymes classification and propertiesorne555Belum ada peringkat
- Bioresource Technology: Ankita Juneja, Frank W.R. Chaplen, Ganti S. MurthyDokumen8 halamanBioresource Technology: Ankita Juneja, Frank W.R. Chaplen, Ganti S. MurthyMermaidBelum ada peringkat
- Enzyme Nomenclature 1978: This Edition Is a Revision of the Recommendations (1972) of the IUPAC–IUB Commission on Biochemical Nomenclature, and Has Been Approved for Publication by the Executive Committee of the International Union of BiochemistryDari EverandEnzyme Nomenclature 1978: This Edition Is a Revision of the Recommendations (1972) of the IUPAC–IUB Commission on Biochemical Nomenclature, and Has Been Approved for Publication by the Executive Committee of the International Union of BiochemistryPenilaian: 4 dari 5 bintang4/5 (3)
- N-Acetyltransferase The Practical Consequences of Polymorphic Activity in ManDokumen16 halamanN-Acetyltransferase The Practical Consequences of Polymorphic Activity in ManIsrael GonzalezBelum ada peringkat
- Ni Hms 473092Dokumen23 halamanNi Hms 473092jp fernandezBelum ada peringkat
- Alcaloides MBDokumen9 halamanAlcaloides MBSule Granados DBelum ada peringkat
- Dong 2015Dokumen12 halamanDong 2015DhanushBelum ada peringkat
- Mannervik 1985Dokumen5 halamanMannervik 1985Branka GlišićBelum ada peringkat
- Differential Expression Profiling of The Hepatic Proteome in A Rat Model of Dioxin ResistanceDokumen13 halamanDifferential Expression Profiling of The Hepatic Proteome in A Rat Model of Dioxin Resistancegiasco78Belum ada peringkat
- Structural Studies of Flaa1 From Helicobacter Pylori Reveal The Mechanism For Inverting 4,6-Dehydratase ActivityDokumen8 halamanStructural Studies of Flaa1 From Helicobacter Pylori Reveal The Mechanism For Inverting 4,6-Dehydratase Activityjuan acostaBelum ada peringkat
- PGR Arun 2011 BDokumen5 halamanPGR Arun 2011 Barunprabhu_dhanapalBelum ada peringkat
- Insect Molecular Biology and BiochemistryDari EverandInsect Molecular Biology and BiochemistryPenilaian: 5 dari 5 bintang5/5 (2)
- Food and Chemical Toxicology: Article InfoDokumen7 halamanFood and Chemical Toxicology: Article InfoNathalia MojicaBelum ada peringkat
- 2011 VERSÃO PUBLICADA Towards A Better Understanding of Ipomoea Asarifolia Toxicity - Evidence of The Involvement of A Leaf LectinDokumen7 halaman2011 VERSÃO PUBLICADA Towards A Better Understanding of Ipomoea Asarifolia Toxicity - Evidence of The Involvement of A Leaf Lectinnilberto2Belum ada peringkat
- Imam A.A. Mekkawy and Fakhr. El-Din LasheinDokumen11 halamanImam A.A. Mekkawy and Fakhr. El-Din Lasheinraeesaa_j09Belum ada peringkat
- CarbohydratDokumen6 halamanCarbohydratVan Anh NguyenBelum ada peringkat
- Metabolic Inhibitors V1: A Comprehensive TreatiseDari EverandMetabolic Inhibitors V1: A Comprehensive TreatiseR HochsterBelum ada peringkat
- The Putative Ovarian Tumour Marker Gene HE4 (WFDC2), Is Expressed in Normal Tissues and Undergoes Complex Alternative Splicing To Yield Multiple Protein IsoformsDokumen6 halamanThe Putative Ovarian Tumour Marker Gene HE4 (WFDC2), Is Expressed in Normal Tissues and Undergoes Complex Alternative Splicing To Yield Multiple Protein IsoformsHanny TanzilBelum ada peringkat
- Sheffield 2000Dokumen8 halamanSheffield 2000laciyeg352Belum ada peringkat
- Genomic Analysis of MAP Kinase Cascades In: Arabidopsis Defense ResponsesDokumen13 halamanGenomic Analysis of MAP Kinase Cascades In: Arabidopsis Defense ResponsesMelvin PrasadBelum ada peringkat
- Effect of Nonylphenol On Male Reproduction Analysis of Rat Epididymal Biochemical Markers and Antioxidant Defense EnzymesDokumen8 halamanEffect of Nonylphenol On Male Reproduction Analysis of Rat Epididymal Biochemical Markers and Antioxidant Defense EnzymesOrlando Karim Shiro Jr.Belum ada peringkat
- An Autoinhibitory Domain Confers Redox Regulation To Maize Glycerate KinaseDokumen9 halamanAn Autoinhibitory Domain Confers Redox Regulation To Maize Glycerate Kinase10sgBelum ada peringkat
- Comparative Genomics of The HOG-signalling System in Fungi: Ó Springer-Verlag 2006Dokumen15 halamanComparative Genomics of The HOG-signalling System in Fungi: Ó Springer-Verlag 2006damien333Belum ada peringkat
- 016 Scas Abstracts Listed Alphabetically by Abstractspdf 19 2016 Scas AbstractsDokumen25 halaman016 Scas Abstracts Listed Alphabetically by Abstractspdf 19 2016 Scas AbstractsdssagBelum ada peringkat
- Bioquímica Lectura 2Dokumen12 halamanBioquímica Lectura 2Paulo MellaBelum ada peringkat
- tmp1589 TMPDokumen18 halamantmp1589 TMPFrontiersBelum ada peringkat
- Interactions of antimalarial drug with lipid membranesDokumen11 halamanInteractions of antimalarial drug with lipid membranesFernanda Stuani PereiraBelum ada peringkat
- Metabolism of Halophilic Archaea Compared Across Four Genome SequencesDokumen20 halamanMetabolism of Halophilic Archaea Compared Across Four Genome SequencesUlfa MutiaraBelum ada peringkat
- Lipases in LipolysisDokumen13 halamanLipases in LipolysisAnonymous wbRUEuDBelum ada peringkat
- International Rice Research Notes Vol.28 No.1Dokumen80 halamanInternational Rice Research Notes Vol.28 No.1ccquintosBelum ada peringkat
- Imperial College MapDokumen3 halamanImperial College MapRuth Alma CotrutaBelum ada peringkat
- Quinque pillars of evolution in the Indian pharmaceutical sectorDokumen99 halamanQuinque pillars of evolution in the Indian pharmaceutical sectorratithaneBelum ada peringkat
- Principles of Inheritance N Variation PDFDokumen4 halamanPrinciples of Inheritance N Variation PDFshodhan shettyBelum ada peringkat
- Microbial Genomes !Dokumen15 halamanMicrobial Genomes !talha153Belum ada peringkat
- Thermal Resistance of Saccharomyces Yeast Ascospores in BeersDokumen6 halamanThermal Resistance of Saccharomyces Yeast Ascospores in BeersIonuț NeaguBelum ada peringkat
- Pro Top Last FusionDokumen13 halamanPro Top Last Fusionj131089Belum ada peringkat
- Cleaning Validation For Biopharmaceutical Manufacturing at GenentechDokumen4 halamanCleaning Validation For Biopharmaceutical Manufacturing at GenentechLisa Denysa SigarBelum ada peringkat
- 6476 Formulation of Alternative Culture Media For Bacterial and Fungal GrowthDokumen6 halaman6476 Formulation of Alternative Culture Media For Bacterial and Fungal GrowthHanief MitsukhaBelum ada peringkat
- (Advances in Biochemical Engineering Biotechnology) K.E.L. Eriksson-Biotechnology in The Pulp and Paper Industry-Springer Verlag (19Dokumen335 halaman(Advances in Biochemical Engineering Biotechnology) K.E.L. Eriksson-Biotechnology in The Pulp and Paper Industry-Springer Verlag (19Josepepe Bedolla BBelum ada peringkat
- Polymerase Chain ReactionDokumen20 halamanPolymerase Chain ReactionGuadalupe Berber100% (1)
- Stomatal Variation in Dicot and Monocots PDFDokumen5 halamanStomatal Variation in Dicot and Monocots PDFReinaur Signey AluningBelum ada peringkat
- 9A23605 ImmunologyDokumen4 halaman9A23605 ImmunologysivabharathamurthyBelum ada peringkat
- Pharma Companies by StateDokumen8 halamanPharma Companies by StateMuhammad Farhan KhanBelum ada peringkat
- The Polycomb Group Protein Yaf2 Regulates The Pluripotency of Embryonic Stem Cells in A Phosphorylation-Dependent MannerDokumen13 halamanThe Polycomb Group Protein Yaf2 Regulates The Pluripotency of Embryonic Stem Cells in A Phosphorylation-Dependent MannerSHUMETBelum ada peringkat
- Time Table Fall 2023 - (Updatded 11-10-2023)Dokumen2 halamanTime Table Fall 2023 - (Updatded 11-10-2023)Riya HereBelum ada peringkat
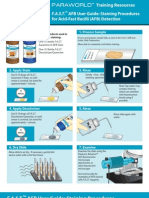
- Fast Userguide StainDokumen2 halamanFast Userguide StainRakesh BagrajiBelum ada peringkat
- Golden Rice Genetic PresentationDokumen23 halamanGolden Rice Genetic PresentationPeter FadoulBelum ada peringkat
- Bigdye Terminator Protocol v1.1Dokumen74 halamanBigdye Terminator Protocol v1.1Citrawati Dyah Kencono WunguBelum ada peringkat
- Handbook of Fungal Biotechnology Vol20Dokumen569 halamanHandbook of Fungal Biotechnology Vol20Trung Huynh100% (1)
- Klasifikasikan Berdasarkan Asalnya Stem CellDokumen2 halamanKlasifikasikan Berdasarkan Asalnya Stem CellfathinanisaBelum ada peringkat
- Acid Phosphatase - Wikipedia, The Free EncyclopediaDokumen3 halamanAcid Phosphatase - Wikipedia, The Free EncyclopediaBinu GeorgeBelum ada peringkat
- Joe Batson and The Scripps Research InstituteDokumen1 halamanJoe Batson and The Scripps Research InstituteJoe BatsonBelum ada peringkat
- Biology SeedsDokumen493 halamanBiology Seedspink921125Belum ada peringkat
- Bryson Ucl Projects2017Dokumen10 halamanBryson Ucl Projects2017DevendraReddyPoreddyBelum ada peringkat
- Levine H-Challenges in The CGMP Manufacturing of Hescs-Lessons Learned From Monoclonal Antibodies-RprintDokumen24 halamanLevine H-Challenges in The CGMP Manufacturing of Hescs-Lessons Learned From Monoclonal Antibodies-RprintChilliG35Belum ada peringkat
- Pullulan Mini Review AMBDokumen18 halamanPullulan Mini Review AMBMohammadbasha MakandarBelum ada peringkat
- Regulation of Gene ExpressionDokumen8 halamanRegulation of Gene ExpressionKevin ApodacaBelum ada peringkat
- Diagnostic Center Feasibility Study - IndiaDokumen33 halamanDiagnostic Center Feasibility Study - IndiaPrafull Hippargekar100% (3)
- AmelogeninaDokumen7 halamanAmelogeninaCelta Gómez TrejoBelum ada peringkat