Anda mungkin juga menyukai
- Candidiasis (Hasznalt)Dokumen4 halamanCandidiasis (Hasznalt)Ilyes FerenczBelum ada peringkat
- Viral Vistas: Insights into Infectious Diseases: The Invisible War: Decoding the Game of Hide and Seek with PathogensDari EverandViral Vistas: Insights into Infectious Diseases: The Invisible War: Decoding the Game of Hide and Seek with PathogensBelum ada peringkat
- Pathophysiology of Oral CandidiasisDokumen11 halamanPathophysiology of Oral CandidiasisAldyTriawanBelum ada peringkat
- Opportunistic Yeast InfectionDokumen14 halamanOpportunistic Yeast Infectionelsa_imamBelum ada peringkat
- Pathogenesis and Treatment of Oral CandidosisDokumen20 halamanPathogenesis and Treatment of Oral CandidosiselisabethpriskaBelum ada peringkat
- Candidiasis (Brut)Dokumen6 halamanCandidiasis (Brut)Ilyes FerenczBelum ada peringkat
- Red and White Lesions of The Oral Mucosa: DR - Wisam RasoolDokumen43 halamanRed and White Lesions of The Oral Mucosa: DR - Wisam RasoolFlorida ManBelum ada peringkat
- Jurnal Englis 2 KtiDokumen17 halamanJurnal Englis 2 KtigilangBelum ada peringkat
- Chap 2Dokumen13 halamanChap 2Aamna HanifBelum ada peringkat
- Shweta PPT ThesisDokumen10 halamanShweta PPT ThesisAKSHAY WAKTEBelum ada peringkat
- Oral Candidiasis: Etiology and PathogenesisDokumen5 halamanOral Candidiasis: Etiology and PathogenesisNovita Indah PratiwiBelum ada peringkat
- CandidiasisDokumen51 halamanCandidiasisLincy JohnyBelum ada peringkat
- CandidaDokumen33 halamanCandidaAlberto ValenzuelaBelum ada peringkat
- Oral Candidiasis - ReviewDokumen9 halamanOral Candidiasis - ReviewAdhelia Galuh PrmtsrBelum ada peringkat
- Management and Treatment of Fungal Infection in The Oral CavityDokumen4 halamanManagement and Treatment of Fungal Infection in The Oral CavityKIU PUBLICATION AND EXTENSIONBelum ada peringkat
- Oral CandidiasisDokumen28 halamanOral CandidiasisjnrrszrnvmBelum ada peringkat
- b.8 Oral CandidiasisDokumen14 halamanb.8 Oral Candidiasisbryamjbriceno100% (1)
- Article WMC004489Dokumen8 halamanArticle WMC004489dyahayueBelum ada peringkat
- Chronic Hyperplastic Candidiasis of The Oral Mucosa Case Report PDFDokumen3 halamanChronic Hyperplastic Candidiasis of The Oral Mucosa Case Report PDFGita Dwi DesianiBelum ada peringkat
- Maxillofacial MicrobiologyDokumen60 halamanMaxillofacial MicrobiologyAbel AbrahamBelum ada peringkat
- Opportunistic MycosesDokumen28 halamanOpportunistic MycosesSally ElhadadBelum ada peringkat
- Opportunistic MycosesDokumen31 halamanOpportunistic MycosesMaxamed Faarax XaashiBelum ada peringkat
- Helicobacter is-WPS OfficeDokumen2 halamanHelicobacter is-WPS Officegeorginho kingsBelum ada peringkat
- Haider OralllllllllllllllllllllllDokumen13 halamanHaider OralllllllllllllllllllllllAli YehyaBelum ada peringkat
- Patho A 1. 11 Infectious Diseases Tagayuna 2015Dokumen10 halamanPatho A 1. 11 Infectious Diseases Tagayuna 2015Ala'a Emerald AguamBelum ada peringkat
- White and Red Lesion Lecture 1 - DR Ameena Ryhan DiajilDokumen43 halamanWhite and Red Lesion Lecture 1 - DR Ameena Ryhan Diajilفاطمة احمد غضبان الزياديBelum ada peringkat
- CandidiasisDokumen19 halamanCandidiasisTeguh Adi PartamaBelum ada peringkat
- Oral Health in America A Report of The Surgeon General (Prosiding)Dokumen4 halamanOral Health in America A Report of The Surgeon General (Prosiding)Zkdlin SpaceBelum ada peringkat
- Tugas Kukel WulanDokumen15 halamanTugas Kukel WulanDonny Ardika NovanandaBelum ada peringkat
- Periodontology 2000 - 2019 - Chen - Herpesvirus Bacteria Synergistic Interaction in PeriodontitisDokumen23 halamanPeriodontology 2000 - 2019 - Chen - Herpesvirus Bacteria Synergistic Interaction in PeriodontitisAdil KaukabBelum ada peringkat
- Candidosis A New ChallengeDokumen7 halamanCandidosis A New ChallengeSalvador contreras huertaBelum ada peringkat
- Review Article: What Makes Oral Candidiasis Recurrent Infection? A Clinical ViewDokumen6 halamanReview Article: What Makes Oral Candidiasis Recurrent Infection? A Clinical ViewAF KoasBelum ada peringkat
- Candidiasis - Red and White Manifestations in The Oral CavityDokumen8 halamanCandidiasis - Red and White Manifestations in The Oral CavityFajarRahmantiyoBelum ada peringkat
- Candidiasis Red and WhiteDokumen8 halamanCandidiasis Red and WhitejnrrszrnvmBelum ada peringkat
- Candidiasis Red and White Manifestations in The Oral CavityDokumen8 halamanCandidiasis Red and White Manifestations in The Oral CavityDaniela Juárez CastellanosBelum ada peringkat
- Winter 1Dokumen6 halamanWinter 1Jam Knows RightBelum ada peringkat
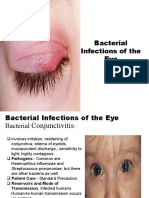
- Bacterial Infections of The EyeDokumen14 halamanBacterial Infections of The EyeThomas GealonBelum ada peringkat
- CandidiasisDokumen3 halamanCandidiasisYanna Habib-MangotaraBelum ada peringkat
- Candidiasis A Fungal Infection Current Challenges and ProgressDokumen11 halamanCandidiasis A Fungal Infection Current Challenges and ProgressLau's WorldBelum ada peringkat
- Candidiasis: International Class MakalahDokumen16 halamanCandidiasis: International Class MakalahIrwan AzizBelum ada peringkat
- GastroDokumen77 halamanGastroMarielle RamosBelum ada peringkat
- Angular Chelitis - StatPearls - NCBI BookshelfDokumen15 halamanAngular Chelitis - StatPearls - NCBI Bookshelfmichaelarchangelus7Belum ada peringkat
- MENINGOCOCCEMIADokumen6 halamanMENINGOCOCCEMIAnikki_villotaBelum ada peringkat
- What Is DiabetesDokumen8 halamanWhat Is DiabetesJessica GintingBelum ada peringkat
- CampyDokumen19 halamanCampyMaria PavlovaBelum ada peringkat
- Oral Candidiasis: Madhu Priya.MDokumen4 halamanOral Candidiasis: Madhu Priya.MinventionjournalsBelum ada peringkat
- Deep Neck and Odontogenic Infections: Key PointsDokumen12 halamanDeep Neck and Odontogenic Infections: Key Pointsnh2411Belum ada peringkat
- Attachment 1Dokumen10 halamanAttachment 1Ara MariBelum ada peringkat
- Intestinal Coccidian: An Overview Epidemiologic Worldwide and ColombiaDokumen14 halamanIntestinal Coccidian: An Overview Epidemiologic Worldwide and ColombiaMARIA LUCIA PIESCHACÓN CHUZCANOBelum ada peringkat
- Clinical Case Reports - 2023 - Lee - Reactivated Herpetic Gingivostomatitis With Secondary Herpes Associated ErythemaDokumen7 halamanClinical Case Reports - 2023 - Lee - Reactivated Herpetic Gingivostomatitis With Secondary Herpes Associated ErythemaAhmad AbrorBelum ada peringkat
- Admin, Artikel 7Dokumen5 halamanAdmin, Artikel 7Aristya SidhartaBelum ada peringkat
- Candidiasis 3Dokumen23 halamanCandidiasis 3ELSA NOVANTIBelum ada peringkat
- Oral CandidasisDokumen39 halamanOral CandidasisArmada Eka FredianBelum ada peringkat
- Mycoses Alimentary: of TheDokumen6 halamanMycoses Alimentary: of TheMouhamed YounousseBelum ada peringkat
- Etiology of Gingivitis: September 2011Dokumen19 halamanEtiology of Gingivitis: September 2011Phúc BùiBelum ada peringkat
- Candida Biofilms and Their Role in Infection: L. Julia DouglasDokumen7 halamanCandida Biofilms and Their Role in Infection: L. Julia DouglasksanbtBelum ada peringkat
- DR A. Di Salvo: CANDIDIASIS (Candida Albicans)Dokumen4 halamanDR A. Di Salvo: CANDIDIASIS (Candida Albicans)Hendris CitraBelum ada peringkat
- Introduction To Medical MicrobiologyDokumen9 halamanIntroduction To Medical MicrobiologyIsba Shadai Estrada GarciaBelum ada peringkat
- Microbiology 1Dokumen57 halamanMicrobiology 1Kyla RamonesBelum ada peringkat
- IvpbhandoutDokumen2 halamanIvpbhandoutapi-283303129Belum ada peringkat
- Seizure: Eugen Trinka, Reetta KälviäinenDokumen9 halamanSeizure: Eugen Trinka, Reetta KälviäinenAaron Nureña JaraBelum ada peringkat
- Orchid Disease, Petal or Blossom Blight PDFDokumen3 halamanOrchid Disease, Petal or Blossom Blight PDFViandi DiniBelum ada peringkat
- MCN - Milestones For Fetal Growth and DevelopmentDokumen5 halamanMCN - Milestones For Fetal Growth and DevelopmentBenjo Dela CruzBelum ada peringkat
- Anesthesia For in Vitro Fertilization: Review ArticleDokumen11 halamanAnesthesia For in Vitro Fertilization: Review ArticleSumangala MagiBelum ada peringkat
- Fetal Biophysical ProfileDokumen17 halamanFetal Biophysical ProfileBangun PurwakaBelum ada peringkat
- 2404-Article Text-7186-1-10-20190315 PDFDokumen7 halaman2404-Article Text-7186-1-10-20190315 PDFnelisaBelum ada peringkat
- 11.21 - Maternal Health Status in The PhilippinesDokumen7 halaman11.21 - Maternal Health Status in The PhilippinesJøshua CruzBelum ada peringkat
- Neurology II 6.04 Traumatic Brain Injury Dr. TanDokumen10 halamanNeurology II 6.04 Traumatic Brain Injury Dr. TanAbi IgsBelum ada peringkat
- Sore ThroatDokumen10 halamanSore Throatapi-551073862Belum ada peringkat
- RCSI Bahrain ConnectED Newsletter Winter 2022-23Dokumen32 halamanRCSI Bahrain ConnectED Newsletter Winter 2022-23Yousif YousifBelum ada peringkat
- Ulcerative ColitisDokumen30 halamanUlcerative ColitisAndika SulistianBelum ada peringkat
- Genetic Analysis of Rare Disease: A Bioinformatic ApproachDokumen55 halamanGenetic Analysis of Rare Disease: A Bioinformatic Approachparetini01Belum ada peringkat
- Surgical Site Infection Investigation Tool: AHRQ Safety Program For SurgeryDokumen5 halamanSurgical Site Infection Investigation Tool: AHRQ Safety Program For SurgeryDewi ListyoriniBelum ada peringkat
- Digital Atlas of Cystoscopy: Jan SchönebeckDokumen48 halamanDigital Atlas of Cystoscopy: Jan SchönebeckdemisBelum ada peringkat
- SOSCG 6jul2013 v5Dokumen32 halamanSOSCG 6jul2013 v5Suhazeli Abdullah100% (2)
- Wildlife Fact File - Mammals, Pgs. 111-120Dokumen20 halamanWildlife Fact File - Mammals, Pgs. 111-120ClearMind84100% (2)
- The Radiology AssistantDokumen30 halamanThe Radiology AssistantIcha IchaBelum ada peringkat
- Fundamentals of Nursing10-19Dokumen19 halamanFundamentals of Nursing10-19escolarBelum ada peringkat
- Reverse TsaDokumen4 halamanReverse Tsaprwill00Belum ada peringkat
- Dental ImplantsDokumen1 halamanDental ImplantsPaWan VelagaBelum ada peringkat
- Clinical Presentation of Abdominal TuberculosisDokumen4 halamanClinical Presentation of Abdominal TuberculosisRizky AmaliahBelum ada peringkat
- Lesch-Nyhan SyndromeDokumen4 halamanLesch-Nyhan Syndromeapi-209017848Belum ada peringkat
- Head Trauma Brain InjuryDokumen54 halamanHead Trauma Brain Injurymirzabb1Belum ada peringkat
- DR Lie Khie Chen - Management of MDR Infection in Sepsis Jade 2016Dokumen33 halamanDR Lie Khie Chen - Management of MDR Infection in Sepsis Jade 2016Astria PermanaBelum ada peringkat
- Complication of Enteral Nutrition PDFDokumen3 halamanComplication of Enteral Nutrition PDFIndra WijayaBelum ada peringkat
- B1001A Practice Final Exam 2023Dokumen15 halamanB1001A Practice Final Exam 2023yeBelum ada peringkat
- 2019 ACC AHA Guidelines For Primary Prevention of CV DiseaseDokumen39 halaman2019 ACC AHA Guidelines For Primary Prevention of CV DiseaseArdel RomeroBelum ada peringkat
- Stroke Signs and Symptoms - CDC - GovDokumen2 halamanStroke Signs and Symptoms - CDC - Govjeanne_mbBelum ada peringkat
- CVBD Easy-To-Digest No 4 BabesiosisDokumen12 halamanCVBD Easy-To-Digest No 4 BabesiosisDocBelum ada peringkat
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Dari EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Penilaian: 3 dari 5 bintang3/5 (1)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedDari EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedPenilaian: 4.5 dari 5 bintang4.5/5 (82)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDDari EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDPenilaian: 5 dari 5 bintang5/5 (3)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDari EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionPenilaian: 4 dari 5 bintang4/5 (404)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDari EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityPenilaian: 4 dari 5 bintang4/5 (32)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDari EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BePenilaian: 2 dari 5 bintang2/5 (1)
- Manipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesDari EverandManipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesPenilaian: 4.5 dari 5 bintang4.5/5 (1412)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsDari EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsBelum ada peringkat
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsDari EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsPenilaian: 4 dari 5 bintang4/5 (4)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsDari EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsPenilaian: 5 dari 5 bintang5/5 (1)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisDari EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisPenilaian: 4.5 dari 5 bintang4.5/5 (42)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaDari EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaPenilaian: 4.5 dari 5 bintang4.5/5 (266)
- The Comfort of Crows: A Backyard YearDari EverandThe Comfort of Crows: A Backyard YearPenilaian: 4.5 dari 5 bintang4.5/5 (23)
- The Obesity Code: Unlocking the Secrets of Weight LossDari EverandThe Obesity Code: Unlocking the Secrets of Weight LossPenilaian: 4 dari 5 bintang4/5 (6)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDari EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityPenilaian: 4.5 dari 5 bintang4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDari EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisPenilaian: 3.5 dari 5 bintang3.5/5 (2)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeDari EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifePenilaian: 4.5 dari 5 bintang4.5/5 (254)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Dari EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Penilaian: 4.5 dari 5 bintang4.5/5 (110)
- To Explain the World: The Discovery of Modern ScienceDari EverandTo Explain the World: The Discovery of Modern SciencePenilaian: 3.5 dari 5 bintang3.5/5 (51)
- Critical Thinking: How to Effectively Reason, Understand Irrationality, and Make Better DecisionsDari EverandCritical Thinking: How to Effectively Reason, Understand Irrationality, and Make Better DecisionsPenilaian: 4.5 dari 5 bintang4.5/5 (39)
- The Marshmallow Test: Mastering Self-ControlDari EverandThe Marshmallow Test: Mastering Self-ControlPenilaian: 4.5 dari 5 bintang4.5/5 (60)
- Dark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingDari EverandDark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingPenilaian: 4 dari 5 bintang4/5 (1138)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessDari EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessPenilaian: 4.5 dari 5 bintang4.5/5 (328)