Anda mungkin juga menyukai
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- High-Level Production of Poly (3-Hydroxybutyrate-co-3-Hydroxyvalerate) by Fed-Batch Culture of Recombinant Escherichia ColiDokumen6 halamanHigh-Level Production of Poly (3-Hydroxybutyrate-co-3-Hydroxyvalerate) by Fed-Batch Culture of Recombinant Escherichia Coliapi-3743140Belum ada peringkat
- 19Dokumen8 halaman19api-3743140Belum ada peringkat
- Production of Aldehyde Oxidases by Microorganisms and Their Enzymatic PropertiesDokumen6 halamanProduction of Aldehyde Oxidases by Microorganisms and Their Enzymatic Propertiesapi-3743140Belum ada peringkat
- Noäi Dung OânDokumen3 halamanNoäi Dung Oânapi-3743140Belum ada peringkat
- Relationships Between Arginine Degradation, PH and Survival in Lactobacillus SakeiDokumen8 halamanRelationships Between Arginine Degradation, PH and Survival in Lactobacillus Sakeiapi-3743140Belum ada peringkat
- Isolation of Nisin-Producing Lactococcus Lactis WNC 20 Strain From Nham, A Traditional Thai Fermented SausageDokumen9 halamanIsolation of Nisin-Producing Lactococcus Lactis WNC 20 Strain From Nham, A Traditional Thai Fermented Sausageapi-3743140Belum ada peringkat
- Competitiveness and Bacteriocin Production of Enterococci in The Production of Spanish-Style Dry Fermented SausagesDokumen10 halamanCompetitiveness and Bacteriocin Production of Enterococci in The Production of Spanish-Style Dry Fermented Sausagesapi-3743140Belum ada peringkat
- ArtDokumen9 halamanArtDaisy Johana Pérez MancipeBelum ada peringkat
- 8Dokumen7 halaman8api-3743140Belum ada peringkat
- Characterization of Lactococci Isolated From Minimally Processed Fresh Fruit and VegetablesDokumen8 halamanCharacterization of Lactococci Isolated From Minimally Processed Fresh Fruit and Vegetablesapi-3743140Belum ada peringkat
- Screening of Lactic Acid Bacteria From Brazilian Meats For Bacteriocin FormationDokumen4 halamanScreening of Lactic Acid Bacteria From Brazilian Meats For Bacteriocin Formationapi-3743140Belum ada peringkat
- 11Dokumen7 halaman11api-3743140Belum ada peringkat
- 7Dokumen10 halaman7api-3743140Belum ada peringkat
- 3Dokumen10 halaman3api-3743140Belum ada peringkat
- Batch and Fed-Batch Production of Butyric Acid by Clostridium Butyricum ZJUCBDokumen5 halamanBatch and Fed-Batch Production of Butyric Acid by Clostridium Butyricum ZJUCBapi-3743140Belum ada peringkat
- 6Dokumen9 halaman6api-3743140100% (1)
- 27Dokumen4 halaman27api-3743140Belum ada peringkat
- 2Dokumen12 halaman2api-3743140100% (1)
- 23Dokumen11 halaman23api-3743140Belum ada peringkat
- 21Dokumen10 halaman21api-3743140Belum ada peringkat
- Tyrosine Decarboxylase Activity of Lactobacillus Brevis IOEB 9809 Isolated From Wine and L. Brevis ATCC 367Dokumen6 halamanTyrosine Decarboxylase Activity of Lactobacillus Brevis IOEB 9809 Isolated From Wine and L. Brevis ATCC 367api-3743140Belum ada peringkat
- 26Dokumen8 halaman26api-3743140100% (1)
- Production of Poly (3-Hydroxybutyrate) by Fed-Batch Culture of Filamentation-Suppressed Recombinant Escherichia ColiDokumen5 halamanProduction of Poly (3-Hydroxybutyrate) by Fed-Batch Culture of Filamentation-Suppressed Recombinant Escherichia Coliapi-3743140Belum ada peringkat
- 16Dokumen13 halaman16api-3743140Belum ada peringkat
- Biodegradation of Formaldehyde Under Anaerobic Conditions: F. Omil, D. Me Ndez, G. Vidal, R. Me Ndez, and J. M. LemaDokumen8 halamanBiodegradation of Formaldehyde Under Anaerobic Conditions: F. Omil, D. Me Ndez, G. Vidal, R. Me Ndez, and J. M. Lemaapi-3743140Belum ada peringkat
- Bacteriocin Production With Lactobacillus Amylovorus DCE 471 Is Improved and Stabilized by Fed-Batch FermentationDokumen8 halamanBacteriocin Production With Lactobacillus Amylovorus DCE 471 Is Improved and Stabilized by Fed-Batch Fermentationapi-3743140Belum ada peringkat
- Production of Poly (3-Hydroxybutyrate) by Fed-Batch Culture of Recombinant Escherichia Coli With A Highly Concentrated Whey SolutionDokumen4 halamanProduction of Poly (3-Hydroxybutyrate) by Fed-Batch Culture of Recombinant Escherichia Coli With A Highly Concentrated Whey Solutionapi-3743140Belum ada peringkat
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (890)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- Five Factors of CleaningDokumen2 halamanFive Factors of CleaningKimberly Bruce De CastroBelum ada peringkat
- Kalayaan Elementary SchoolDokumen3 halamanKalayaan Elementary SchoolEmmanuel MejiaBelum ada peringkat
- Reliability EngineeringDokumen9 halamanReliability Engineeringnvaradharajan1971Belum ada peringkat
- MTech Information Security FINAL 10052018Dokumen20 halamanMTech Information Security FINAL 10052018sirisha vBelum ada peringkat
- VLSI Design: Introduction & Motivation Introduction & MotivationDokumen33 halamanVLSI Design: Introduction & Motivation Introduction & MotivationPriyanka SharmaBelum ada peringkat
- ABC-S Plus Web Info '08Dokumen4 halamanABC-S Plus Web Info '08anon-331232Belum ada peringkat
- Enzyme Inhibition and ToxicityDokumen12 halamanEnzyme Inhibition and ToxicityDaniel OmolewaBelum ada peringkat
- QT140 500 KG Per Hr. Fish Feed Pelleting PlantDokumen11 halamanQT140 500 KG Per Hr. Fish Feed Pelleting PlantShekhar MitraBelum ada peringkat
- QAF10A200S TheTimkenCompany 2DSalesDrawing 03 06 2023Dokumen1 halamanQAF10A200S TheTimkenCompany 2DSalesDrawing 03 06 2023LeroyBelum ada peringkat
- PM Master Data Template v1Dokumen72 halamanPM Master Data Template v1Naseer SultanBelum ada peringkat
- Python Programming Lecture#2 - Functions, Lists, Packages & Formatting I/ODokumen69 halamanPython Programming Lecture#2 - Functions, Lists, Packages & Formatting I/OHamsa VeniBelum ada peringkat
- Bellin, E. H. (1984) - The Psychoanalytic Narrative On The Transformational Axis Between Writing and SpeechDokumen15 halamanBellin, E. H. (1984) - The Psychoanalytic Narrative On The Transformational Axis Between Writing and SpeechofanimenochBelum ada peringkat
- Measurements/ Specifications: Torque Wrench Selection GuideDokumen5 halamanMeasurements/ Specifications: Torque Wrench Selection GuideSylvester RakgateBelum ada peringkat
- DbintfcDokumen80 halamanDbintfchnr.uninstallBelum ada peringkat
- UniversulDokumen56 halamanUniversulGabriel DutuBelum ada peringkat
- Transportation Planning ProcessDokumen43 halamanTransportation Planning ProcessAncheta Suzanne ClarisseBelum ada peringkat
- Valve Actuator Data SheetDokumen1 halamanValve Actuator Data SheetRob StorrowBelum ada peringkat
- UnderstandingCryptology CoreConcepts 6-2-2013Dokumen128 halamanUnderstandingCryptology CoreConcepts 6-2-2013zenzei_Belum ada peringkat
- Metacentric Height: From Wikipedia, The Free EncyclopediaDokumen6 halamanMetacentric Height: From Wikipedia, The Free EncyclopediaВладимир ШевченкоBelum ada peringkat
- Hydrogen and Its Compound.1Dokumen10 halamanHydrogen and Its Compound.1abhishekBelum ada peringkat
- Solution of Homework 4Dokumen4 halamanSolution of Homework 4Kamalakar Reddy100% (2)
- Haidarali-MR-2011-PhD-Thesis 01 PDFDokumen378 halamanHaidarali-MR-2011-PhD-Thesis 01 PDFIbrahim KhanBelum ada peringkat
- Grade 5 Mtap Compilation 2011,2013 2015 2016 2017 Division OralsDokumen5 halamanGrade 5 Mtap Compilation 2011,2013 2015 2016 2017 Division OralsChristine De San JoseBelum ada peringkat
- 2 To 12F SM UT Armoured OFC-2Steel Wire Design Sheet-7.5 MMDokumen3 halaman2 To 12F SM UT Armoured OFC-2Steel Wire Design Sheet-7.5 MMTropic BazarBelum ada peringkat
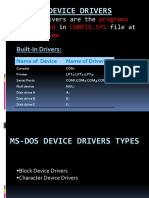
- Ms-Dos Device Drivers: Device Drivers Are The That in File atDokumen13 halamanMs-Dos Device Drivers: Device Drivers Are The That in File atJass GillBelum ada peringkat
- Service Manual: S&T Motors Co., LTDDokumen94 halamanService Manual: S&T Motors Co., LTDJuliano PedrosoBelum ada peringkat
- Simultaneous EquationsDokumen11 halamanSimultaneous EquationsSaleena AurangzaibBelum ada peringkat
- WPS Office 2016Dokumen22 halamanWPS Office 2016Muhammad Aliff SyukriBelum ada peringkat
- Toyo ML210 thread cutting gear assembly drawingsDokumen12 halamanToyo ML210 thread cutting gear assembly drawingsiril anwarBelum ada peringkat
- Rso PDFDokumen120 halamanRso PDFjohn shepardBelum ada peringkat