Anda mungkin juga menyukai
- Mecanismos Que Regulan La Actividad Del Receptor de Quimiocinas 2011Dokumen11 halamanMecanismos Que Regulan La Actividad Del Receptor de Quimiocinas 2011oscarbio2009Belum ada peringkat
- Chemical Molecules That Regulate Transcription and Facilitate Cell-to-Cell CommunicationDokumen11 halamanChemical Molecules That Regulate Transcription and Facilitate Cell-to-Cell CommunicationazzaassBelum ada peringkat
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsDari EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneBelum ada peringkat
- Complement and The Regulation of T Cell ResponsesDokumen41 halamanComplement and The Regulation of T Cell ResponsesFrancelia Quiñonez RuvalcabaBelum ada peringkat
- Coagulation and Innate Immune Responses: Can We View Them Separately?Dokumen9 halamanCoagulation and Innate Immune Responses: Can We View Them Separately?nuraswadBelum ada peringkat
- Las Quimiocinas Hacen Sonar La Alarma El Papel de La Quimiocina Atípica en La Inflamación y El Cáncer 2018Dokumen9 halamanLas Quimiocinas Hacen Sonar La Alarma El Papel de La Quimiocina Atípica en La Inflamación y El Cáncer 2018oscarbio2009Belum ada peringkat
- Bacterial Chemoreceptors: High-Performance Signaling in Networked ArraysDokumen11 halamanBacterial Chemoreceptors: High-Performance Signaling in Networked ArrayslazaroczBelum ada peringkat
- Opioids, Opioid Receptors, and The Immune Response: ReviewDokumen13 halamanOpioids, Opioid Receptors, and The Immune Response: ReviewAulas EspañolBelum ada peringkat
- Regulación de La Hematopoyesis Por El Sistema de Quimioquinas 2018Dokumen5 halamanRegulación de La Hematopoyesis Por El Sistema de Quimioquinas 2018oscarbio2009Belum ada peringkat
- 1 s2.0 S1535947621001584 MainDokumen28 halaman1 s2.0 S1535947621001584 MainBen DresimBelum ada peringkat
- At The Crossroads of Lipid Metabolism and InflammationDokumen7 halamanAt The Crossroads of Lipid Metabolism and InflammationChow Sing SingBelum ada peringkat
- Mono Vs PoliDokumen11 halamanMono Vs PoliYondri Mandaku TasidjawaBelum ada peringkat
- Belas Figuras Nada de CampyDokumen6 halamanBelas Figuras Nada de CampyHaurusBelum ada peringkat
- Transcription Sample Old Essay PDFDokumen7 halamanTranscription Sample Old Essay PDFOccamsRazorBelum ada peringkat
- Microreview: Quorum Sensing by Peptide Pheromones and Two-Component Signal-Transduction Systems in Gram-Positive BacteriaDokumen10 halamanMicroreview: Quorum Sensing by Peptide Pheromones and Two-Component Signal-Transduction Systems in Gram-Positive BacteriaCristina AlexandraBelum ada peringkat
- (14796813 - Journal of Molecular Endocrinology) Chromatin Immunoprecipitation - Advancing Analysis of Nuclear Hormone SignalingDokumen11 halaman(14796813 - Journal of Molecular Endocrinology) Chromatin Immunoprecipitation - Advancing Analysis of Nuclear Hormone Signalingderek lauBelum ada peringkat
- Wang 2020 - The NLRP3 Inflammasome - Mechanism of Action, Role in Disease and TherapiesDokumen12 halamanWang 2020 - The NLRP3 Inflammasome - Mechanism of Action, Role in Disease and TherapiesShirley AlvesBelum ada peringkat
- Static and Dynamic Interactions Between Endothelium and Circulating Cells in CancerDokumen25 halamanStatic and Dynamic Interactions Between Endothelium and Circulating Cells in CancerderBelum ada peringkat
- 2a PDFDokumen11 halaman2a PDFAndrea Il MiodoBelum ada peringkat
- High Expression of Oxidative Phosphorylation Genes Predicts Improved Survival in Squamous Cell Carcinomas of The Head and Neck and LungDokumen14 halamanHigh Expression of Oxidative Phosphorylation Genes Predicts Improved Survival in Squamous Cell Carcinomas of The Head and Neck and Lungsalmaa safiraBelum ada peringkat
- Wecb 646Dokumen9 halamanWecb 646azzaassBelum ada peringkat
- Chaperon AsDokumen6 halamanChaperon AsVale GerónimoBelum ada peringkat
- Regulacion Metabolica de La EpigeneticaDokumen9 halamanRegulacion Metabolica de La EpigeneticaDaniel J Alejos CBelum ada peringkat
- Endosomal Pathway Role of TRP Channels in The Regulation of TheDokumen10 halamanEndosomal Pathway Role of TRP Channels in The Regulation of ThealrokkBelum ada peringkat
- Cellular and Molecular Mechanisms of Inflammation: Vascular Adhesion MoleculesDari EverandCellular and Molecular Mechanisms of Inflammation: Vascular Adhesion MoleculesCharles G. CochraneBelum ada peringkat
- Single-Cell Metabolic Profiling of Human Cytotoxic T Cells: ArticlesDokumen19 halamanSingle-Cell Metabolic Profiling of Human Cytotoxic T Cells: ArticlesShivaprakash Jagalur MuttBelum ada peringkat
- Choi2016 1Dokumen14 halamanChoi2016 1Jose Angel AbadíaBelum ada peringkat
- Mammalian Synthetic Biology - From Tools To Therapies: Dominique Aubel, and Martin FusseneggerDokumen14 halamanMammalian Synthetic Biology - From Tools To Therapies: Dominique Aubel, and Martin FusseneggerChickencito PerezBelum ada peringkat
- TMP 662Dokumen11 halamanTMP 662FrontiersBelum ada peringkat
- Tema 3 - Inmune Responses To ImplantsDokumen18 halamanTema 3 - Inmune Responses To ImplantsIsrael GonzálezBelum ada peringkat
- بكتريا PDFDokumen45 halamanبكتريا PDFADEL A. ABDBelum ada peringkat
- Arrowsmith Et Al (2012) - Epigenetic Protein Families A New Frontier For Drug DiscoveryDokumen17 halamanArrowsmith Et Al (2012) - Epigenetic Protein Families A New Frontier For Drug DiscoveryRabiatul AdawiyahBelum ada peringkat
- Tunn A Cliff e Chubba Am 2020Dokumen23 halamanTunn A Cliff e Chubba Am 2020ano3.nyme1415926535Belum ada peringkat
- NF-KB and The Immune ResponseDokumen23 halamanNF-KB and The Immune ResponseAndri Praja SatriaBelum ada peringkat
- Inter-Cellular Nanovesicle Mediated MicroRNA TransferDokumen19 halamanInter-Cellular Nanovesicle Mediated MicroRNA Transferanon_579968442Belum ada peringkat
- Three Dimensional Genome Architecture Players and MechanismsDokumen13 halamanThree Dimensional Genome Architecture Players and MechanismsNatalio Vazquez CruzBelum ada peringkat
- Transduction Protein Interactions Define Specificity in Signal ProteinDokumen22 halamanTransduction Protein Interactions Define Specificity in Signal ProteinnitishpathaniaBelum ada peringkat
- Insights Into The Novel Function of System XC - in Regulated Cell DeathDokumen13 halamanInsights Into The Novel Function of System XC - in Regulated Cell DeathCom DigfulBelum ada peringkat
- The Many Roles of Chemokines and Chemokine Receptors in InflammationDokumen12 halamanThe Many Roles of Chemokines and Chemokine Receptors in InflammationMedranoReyesLuisinBelum ada peringkat
- Mechanism of Action of Hydroxychloroquine As An Antirheumatic DrugDokumen10 halamanMechanism of Action of Hydroxychloroquine As An Antirheumatic DrugJim MorrisonBelum ada peringkat
- Interesting Biology Topics 2023Dokumen10 halamanInteresting Biology Topics 2023Elchin HuseynovBelum ada peringkat
- HHS Public Access: Unraveling The Complex Interplay Between T Cell Metabolism and FunctionDokumen33 halamanHHS Public Access: Unraveling The Complex Interplay Between T Cell Metabolism and Functionro111111Belum ada peringkat
- Advances in Regulatory Network 2011Dokumen15 halamanAdvances in Regulatory Network 2011nungkionBelum ada peringkat
- The Role of Natural Killer Cells in Chronic Myeloid LeukemiaDokumen5 halamanThe Role of Natural Killer Cells in Chronic Myeloid LeukemiaRicardo MeloBelum ada peringkat
- Cell Junctions, Gap Adhesions, and ExtracellularDokumen58 halamanCell Junctions, Gap Adhesions, and ExtracellularFabian Christine MaeBelum ada peringkat
- Metabolomic Profiling of Drug Responses in Acute Myeloid Leukaemia Cell LinesDokumen10 halamanMetabolomic Profiling of Drug Responses in Acute Myeloid Leukaemia Cell LinesGaurav SharmaBelum ada peringkat
- High-Throughput Single Molecule Tracking Identifies Drug Interactions and Cellular MechanismsDokumen34 halamanHigh-Throughput Single Molecule Tracking Identifies Drug Interactions and Cellular MechanismsscrfciolagettBelum ada peringkat
- 2009 Effect Kaqun WaterDokumen12 halaman2009 Effect Kaqun Watermarjan sharabianiBelum ada peringkat
- The Reactome Pathway KnowledgebaseDokumen6 halamanThe Reactome Pathway KnowledgebaseAndressa AlvesBelum ada peringkat
- NIH Public Access: Author ManuscriptDokumen25 halamanNIH Public Access: Author ManuscriptZakia DrajatBelum ada peringkat
- Psychoneuroimmunology: F T, I E & J MDokumen10 halamanPsychoneuroimmunology: F T, I E & J MPauloBelum ada peringkat
- Immunologic Pathomechanism of Hodgkin's Lymphoma: ReviewDokumen10 halamanImmunologic Pathomechanism of Hodgkin's Lymphoma: ReviewPaola Alexandra Chavez FBelum ada peringkat
- Programmed Cell Death in Plants: A Role For Mitochondrial-Associated HexokinasesDokumen3 halamanProgrammed Cell Death in Plants: A Role For Mitochondrial-Associated HexokinasesMimô SABelum ada peringkat
- The Basis For The Immunoregulatory Role of Macrophages and Other Accessory Cells.Dokumen7 halamanThe Basis For The Immunoregulatory Role of Macrophages and Other Accessory Cells.Cristian Gutiérrez VeraBelum ada peringkat
- Research ProposalDokumen7 halamanResearch ProposalBitan BiswasBelum ada peringkat
- 016 Scas Abstracts Listed Alphabetically by Abstractspdf 19 2016 Scas AbstractsDokumen25 halaman016 Scas Abstracts Listed Alphabetically by Abstractspdf 19 2016 Scas AbstractsdssagBelum ada peringkat
- Colwrt 161 Mini-Lit-ReviewDokumen18 halamanColwrt 161 Mini-Lit-Reviewapi-626218608Belum ada peringkat
- Chromosone AnalysisDokumen234 halamanChromosone AnalysisSpataru VasileBelum ada peringkat
- Hanahan and Weinberg, 2000Dokumen6 halamanHanahan and Weinberg, 2000AdityaBelum ada peringkat
- Slides and Notes - ChChemical Signals and Cell Communicationemical Signals and Cell CommunicationDokumen10 halamanSlides and Notes - ChChemical Signals and Cell Communicationemical Signals and Cell CommunicationadyejzinedBelum ada peringkat
- Types of FeverDokumen10 halamanTypes of Fevermd191031Belum ada peringkat
- WK13 CDN - Cholera, Bacillary Dysentery, Amoebiasis, Typhoid Fever, Hepatitis ADokumen38 halamanWK13 CDN - Cholera, Bacillary Dysentery, Amoebiasis, Typhoid Fever, Hepatitis APotato PceeBelum ada peringkat
- BK Case ReportDokumen4 halamanBK Case ReportEngidaBelum ada peringkat
- MGY378 2014 Outline (1) - 88937Dokumen4 halamanMGY378 2014 Outline (1) - 88937Christine QianBelum ada peringkat
- Gillen Bare Sy PDFDokumen6 halamanGillen Bare Sy PDFberinah65Belum ada peringkat
- Health 9 Venereal Diseases: - Caused by A Bacteria Named Chlamydia TrachomatisDokumen8 halamanHealth 9 Venereal Diseases: - Caused by A Bacteria Named Chlamydia TrachomatisClaire MedianaBelum ada peringkat
- Handouts - 6BIOM002W.2 - Inflammation 2018-9 PDFDokumen66 halamanHandouts - 6BIOM002W.2 - Inflammation 2018-9 PDFFatima HusseinBelum ada peringkat
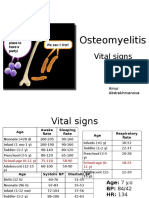
- PBL 2 - OsteomyelitisDokumen10 halamanPBL 2 - OsteomyelitisAinur AbdrakhmanovaBelum ada peringkat
- Indriatmi, W. 2018. Ilmu Penyakit Dan Kulit Kelamin. Edisi Ke-7. Jakarta: Badan Penerbit Fakultas Kedokteran IndonesiaDokumen10 halamanIndriatmi, W. 2018. Ilmu Penyakit Dan Kulit Kelamin. Edisi Ke-7. Jakarta: Badan Penerbit Fakultas Kedokteran IndonesiaDiza Hanni PertiwiBelum ada peringkat
- Fever and RashDokumen33 halamanFever and RashVarun B RenukappaBelum ada peringkat
- WEEK 10 LABORATORY EXERCISE - Lymphatic Immune SystemDokumen7 halamanWEEK 10 LABORATORY EXERCISE - Lymphatic Immune SystemEricka ElloBelum ada peringkat
- Capaian Mutu Respon Time Igd-2Dokumen1.536 halamanCapaian Mutu Respon Time Igd-2Arsyad AkmatulBelum ada peringkat
- Yellow Fever Vaccination Centre in MalaysiaDokumen9 halamanYellow Fever Vaccination Centre in MalaysiaAngeline LeeBelum ada peringkat
- Bacte Sample QuestionDokumen45 halamanBacte Sample QuestionpikachuBelum ada peringkat
- Herpes Brochure PDFDokumen2 halamanHerpes Brochure PDFEuresha JeremiahBelum ada peringkat
- 6 MonocytopoiesisDokumen8 halaman6 MonocytopoiesisVannisa Dwi NovianaBelum ada peringkat
- Managing Allergic Rhinitis in Children Through Ayurvedic Herbal MedicinesDokumen11 halamanManaging Allergic Rhinitis in Children Through Ayurvedic Herbal MedicinesAyu WulandariBelum ada peringkat
- BMS533 Lab PracticalDokumen3 halamanBMS533 Lab PracticalYUSNURDALILA DALIBelum ada peringkat
- Who Influenza SurveillanceDokumen153 halamanWho Influenza SurveillanceRidho Al FiqriBelum ada peringkat
- RSV Respiratory Syncytial Virus: Identification TreatmentDokumen1 halamanRSV Respiratory Syncytial Virus: Identification Treatmentapi-350983586Belum ada peringkat
- Anti Biotic WhoDokumen4 halamanAnti Biotic WhoWelfredo Jr YuBelum ada peringkat
- Celiac Disease in ChildrenDokumen59 halamanCeliac Disease in Childrend-fbuser-57045067Belum ada peringkat
- Asepsis and Infection Control (Week 4)Dokumen7 halamanAsepsis and Infection Control (Week 4)JEREMY MAKALINTALBelum ada peringkat
- AQA GCSE Bio Combined End of Topic B6Dokumen6 halamanAQA GCSE Bio Combined End of Topic B6josephBelum ada peringkat
- CV-polar Flagella PE - Multiple Flagella: Spore Survives in BC GRAM POS OBLIGATE AEROBE - Bacillus, NocardiaDokumen4 halamanCV-polar Flagella PE - Multiple Flagella: Spore Survives in BC GRAM POS OBLIGATE AEROBE - Bacillus, NocardiaMica SaeronBelum ada peringkat
- Allogeneic Stem Cell Transplantation - M. Laughlin, H. Lazarus (Humana, 2003) WWDokumen466 halamanAllogeneic Stem Cell Transplantation - M. Laughlin, H. Lazarus (Humana, 2003) WWSerah LightBelum ada peringkat
- Handbook For Vaccine and Cold Chain HandlersDokumen128 halamanHandbook For Vaccine and Cold Chain HandlersRyza Jazid Baharuddin NurBelum ada peringkat
- Rabies WHODokumen5 halamanRabies WHOReno Jun NagasanBelum ada peringkat
- Revised National TB Control ProgramDokumen12 halamanRevised National TB Control ProgramsaurabhBelum ada peringkat
- Disease As Oracle Anamnesis Diagnosis An PDFDokumen15 halamanDisease As Oracle Anamnesis Diagnosis An PDFHarsh KhiraiyaBelum ada peringkat