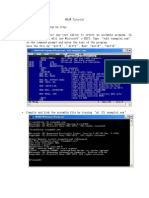
Anda mungkin juga menyukai
- Never Split the Difference: Negotiating As If Your Life Depended On ItDari EverandNever Split the Difference: Negotiating As If Your Life Depended On ItPenilaian: 4.5 dari 5 bintang4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeDari EverandShoe Dog: A Memoir by the Creator of NikePenilaian: 4.5 dari 5 bintang4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDari EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifePenilaian: 4 dari 5 bintang4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)Dari EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Penilaian: 4 dari 5 bintang4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDari EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RacePenilaian: 4 dari 5 bintang4/5 (894)
- The Little Book of Hygge: Danish Secrets to Happy LivingDari EverandThe Little Book of Hygge: Danish Secrets to Happy LivingPenilaian: 3.5 dari 5 bintang3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDari EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FuturePenilaian: 4.5 dari 5 bintang4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDari EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryPenilaian: 3.5 dari 5 bintang3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDari EverandGrit: The Power of Passion and PerseverancePenilaian: 4 dari 5 bintang4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDari EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaPenilaian: 4.5 dari 5 bintang4.5/5 (265)
- On Fire: The (Burning) Case for a Green New DealDari EverandOn Fire: The (Burning) Case for a Green New DealPenilaian: 4 dari 5 bintang4/5 (73)
- The Emperor of All Maladies: A Biography of CancerDari EverandThe Emperor of All Maladies: A Biography of CancerPenilaian: 4.5 dari 5 bintang4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDari EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersPenilaian: 4.5 dari 5 bintang4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaDari EverandThe Unwinding: An Inner History of the New AmericaPenilaian: 4 dari 5 bintang4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnDari EverandTeam of Rivals: The Political Genius of Abraham LincolnPenilaian: 4.5 dari 5 bintang4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDari EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyPenilaian: 3.5 dari 5 bintang3.5/5 (2219)
- Rise of ISIS: A Threat We Can't IgnoreDari EverandRise of ISIS: A Threat We Can't IgnorePenilaian: 3.5 dari 5 bintang3.5/5 (137)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDari EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You ArePenilaian: 4 dari 5 bintang4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Dari EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Penilaian: 4.5 dari 5 bintang4.5/5 (119)
- Her Body and Other Parties: StoriesDari EverandHer Body and Other Parties: StoriesPenilaian: 4 dari 5 bintang4/5 (821)
- The Voice of PLC 1101Dokumen6 halamanThe Voice of PLC 1101The Plymouth Laryngectomy ClubBelum ada peringkat
- The Big Banana by Roberto QuesadaDokumen257 halamanThe Big Banana by Roberto QuesadaArte Público Press100% (2)
- 5.2.1 1539323575 2163Dokumen30 halaman5.2.1 1539323575 2163Brinda TBelum ada peringkat
- HDFDJH 5Dokumen7 halamanHDFDJH 5balamuruganBelum ada peringkat
- Classical Music Forms ExplainedDokumen11 halamanClassical Music Forms Explainedcorinna_harrison100% (1)
- Operations Management Dr. Loay Salhieh Case Study #1: Students: Hadil Mosa Marah Akroush Mohammad Rajab Ousama SammawiDokumen6 halamanOperations Management Dr. Loay Salhieh Case Study #1: Students: Hadil Mosa Marah Akroush Mohammad Rajab Ousama SammawiHadeel Almousa100% (1)
- English 2Dokumen53 halamanEnglish 2momonunu momonunuBelum ada peringkat
- Lecture1 SEODokumen24 halamanLecture1 SEOUsman AnwarBelum ada peringkat
- Safety Data Sheet - en - (68220469) Aluminium Silicate QP (1318!74!7)Dokumen6 halamanSafety Data Sheet - en - (68220469) Aluminium Silicate QP (1318!74!7)sergio.huete.hernandezBelum ada peringkat
- SKILLS TRANSFER PLAN FOR MAINTENANCE OF NAVAL EQUIPMENTDokumen2 halamanSKILLS TRANSFER PLAN FOR MAINTENANCE OF NAVAL EQUIPMENTZaid NordienBelum ada peringkat
- Subsurface Sewage DisposalDokumen174 halamanSubsurface Sewage DisposalSanthi KrishnaBelum ada peringkat
- Mcquillin Murphy ResumeDokumen1 halamanMcquillin Murphy Resumeapi-253430225Belum ada peringkat
- Design and Implementation of Land and Property Ownership Management System in Urban AreasDokumen82 halamanDesign and Implementation of Land and Property Ownership Management System in Urban AreasugochukwuBelum ada peringkat
- BLUEBOOK CITATION GUIDEDokumen12 halamanBLUEBOOK CITATION GUIDEMichaela PortarcosBelum ada peringkat
- V-AMP3: User ManualDokumen19 halamanV-AMP3: User Manualnutmeg_kickerBelum ada peringkat
- Case Study 1Dokumen2 halamanCase Study 1Diana Therese CuadraBelum ada peringkat
- Thompson Industrial Products Inc Is A DiversifiedDokumen4 halamanThompson Industrial Products Inc Is A DiversifiedKailash KumarBelum ada peringkat
- LEGAL STATUs of A PersonDokumen24 halamanLEGAL STATUs of A Personpravas naikBelum ada peringkat
- MASM Tutorial PDFDokumen10 halamanMASM Tutorial PDFShashankDwivediBelum ada peringkat
- PDF. Art Appre - Module 1Dokumen36 halamanPDF. Art Appre - Module 1marvin fajardoBelum ada peringkat
- Alvin - Goldman - and - Dennis - Whitcomb (Eds) - Social - Epistemology - Essential - Readings - 2011 PDFDokumen368 halamanAlvin - Goldman - and - Dennis - Whitcomb (Eds) - Social - Epistemology - Essential - Readings - 2011 PDFOvejaNegra100% (2)
- PSE Inc. V CA G.R. No. 125469, Oct 27, 1997Dokumen7 halamanPSE Inc. V CA G.R. No. 125469, Oct 27, 1997mae ann rodolfoBelum ada peringkat
- Real Estate Marketing Agent Registration Form: Important InstructionsDokumen7 halamanReal Estate Marketing Agent Registration Form: Important InstructionsAshok KumarBelum ada peringkat
- Volatility Clustering, Leverage Effects and Risk-Return Trade-Off in The Nigerian Stock MarketDokumen14 halamanVolatility Clustering, Leverage Effects and Risk-Return Trade-Off in The Nigerian Stock MarketrehanbtariqBelum ada peringkat
- Course Title: Cost Accounting Course Code:441 BBA Program Lecture-3Dokumen20 halamanCourse Title: Cost Accounting Course Code:441 BBA Program Lecture-3Tanvir Ahmed ChowdhuryBelum ada peringkat
- Isha Hatha Yoga - Program Registration FormDokumen2 halamanIsha Hatha Yoga - Program Registration FormKeyur GadaBelum ada peringkat
- CHP - 3 DatabaseDokumen5 halamanCHP - 3 DatabaseNway Nway Wint AungBelum ada peringkat
- Resume John BunkerDokumen8 halamanResume John BunkerJohn BunkerBelum ada peringkat
- SWSP6033 00 2022T3 V1.0-1Dokumen14 halamanSWSP6033 00 2022T3 V1.0-1ayman.abaidallah1990Belum ada peringkat
- Specification For Diesel Engines: AugustDokumen19 halamanSpecification For Diesel Engines: AugustSerge RINAUDOBelum ada peringkat