Anda mungkin juga menyukai
- Fisio Transporte de OxigenoDokumen18 halamanFisio Transporte de OxigenoAndrés MoraBelum ada peringkat
- Transporte de oxígeno y dióxido de carbono en la sangre y tejidosDokumen8 halamanTransporte de oxígeno y dióxido de carbono en la sangre y tejidosTefa 15Belum ada peringkat
- Capítulo 41 Guyton ResumenDokumen5 halamanCapítulo 41 Guyton ResumenJannely Nava MejíaBelum ada peringkat
- Protocolo Ventilacion Pulmonar 1Dokumen20 halamanProtocolo Ventilacion Pulmonar 1Ruth Esther Nuñez MaldonadoBelum ada peringkat
- 40 Transporte Del O2 y Co2 en La Sangre y Los Liquidos Tisulares Guyton 2 8 PDFDokumen7 halaman40 Transporte Del O2 y Co2 en La Sangre y Los Liquidos Tisulares Guyton 2 8 PDFAna RodriguezBelum ada peringkat
- Transporte de O2 y CO2 en La Sangre y Los Líquidos WORDDokumen6 halamanTransporte de O2 y CO2 en La Sangre y Los Líquidos WORDJessica Roman SejasBelum ada peringkat
- Transporte de OxigenoDokumen35 halamanTransporte de OxigenoGabuxita RodriguezBelum ada peringkat
- Trabajo Extra - MergedDokumen26 halamanTrabajo Extra - MergedGeraldyne HernandezBelum ada peringkat
- Capitilo40 Transportedeo2yco2 121128200855 Phpapp02Dokumen42 halamanCapitilo40 Transportedeo2yco2 121128200855 Phpapp02Ginna Paola SaavedraBelum ada peringkat
- Sistema respiratorioDokumen32 halamanSistema respiratorioDaniel Montiel.Belum ada peringkat
- NEUMODokumen35 halamanNEUMOLUZ ASTRID ELIGIO ANAYABelum ada peringkat
- Resumen Capitulo 41 Transporte O2 y Co2Dokumen5 halamanResumen Capitulo 41 Transporte O2 y Co2Lii LiiBelum ada peringkat
- Transporte de Oxigeno de Los Pulmones A Los Tejidos Del OrganismoDokumen5 halamanTransporte de Oxigeno de Los Pulmones A Los Tejidos Del OrganismoEvelyn Calva0% (1)
- Intercambio de OxigenoDokumen22 halamanIntercambio de OxigenoSalvador BARRON MELOBelum ada peringkat
- 004 Transporte de Oxigeno y Dioxido de CarbonoDokumen32 halaman004 Transporte de Oxigeno y Dioxido de CarbonoThulio MoutinhoBelum ada peringkat
- Fisilogia de La RespiracionDokumen5 halamanFisilogia de La RespiracionAlejandro CardozaBelum ada peringkat
- PRÁCTICA 10 Transporte de Gases y Liquidos CorporalesDokumen13 halamanPRÁCTICA 10 Transporte de Gases y Liquidos CorporalesNoemi VelasquezBelum ada peringkat
- TP Mecanismo RespiratorioDokumen6 halamanTP Mecanismo RespiratoriomailenBelum ada peringkat
- Clase Difusión de Gases Pulmonar 2017Dokumen60 halamanClase Difusión de Gases Pulmonar 2017Jose Luis VargasBelum ada peringkat
- Capitulo 41 526237 Downloadable 3079102Dokumen24 halamanCapitulo 41 526237 Downloadable 3079102bbmamani-esBelum ada peringkat
- Tema 3 Introducción Fisiología PsicologíaDokumen5 halamanTema 3 Introducción Fisiología PsicologíaMartaBelum ada peringkat
- Transporte e Intercambio Gaseoso. TextoDokumen7 halamanTransporte e Intercambio Gaseoso. TextoCarolina Rivera ReyBelum ada peringkat
- Reporte de Lectura Capítulo 41 Transporte de Oxígeno y Dioxido de Carbono en La Sangre y Los Líquidos TisularesDokumen8 halamanReporte de Lectura Capítulo 41 Transporte de Oxígeno y Dioxido de Carbono en La Sangre y Los Líquidos TisularesMiguel CórdovaBelum ada peringkat
- Capítulo 40 y 41Dokumen7 halamanCapítulo 40 y 41Wildaury Fondeur RodriguezBelum ada peringkat
- Fisiologia RespiratoriaDokumen60 halamanFisiologia RespiratoriaLesli CortesBelum ada peringkat
- Transporte de oxígeno y dióxido de carbono en la sangreDokumen5 halamanTransporte de oxígeno y dióxido de carbono en la sangreRuth Iraipi QuirogaBelum ada peringkat
- Intercambio de GasesDokumen2 halamanIntercambio de GasesGeraldine AlarconBelum ada peringkat
- Capitulo 41 Transporte de Oxigeno y Dioxido de Carbono en La Sangre y Los Liquidos Tisulares 96163 DownloableDokumen3 halamanCapitulo 41 Transporte de Oxigeno y Dioxido de Carbono en La Sangre y Los Liquidos Tisulares 96163 DownloableGiselle De La CruzBelum ada peringkat
- Alteraciones Del Transporte de OxígenoDokumen7 halamanAlteraciones Del Transporte de OxígenoArely Garrido FloresBelum ada peringkat
- Proceso de Intercambio Alveolo PulmonarDokumen15 halamanProceso de Intercambio Alveolo Pulmonarbssa1965Belum ada peringkat
- Resumen Fisiologia de Guyton Capitulo 40 Ed 12Dokumen6 halamanResumen Fisiologia de Guyton Capitulo 40 Ed 12XDXDXDXDBelum ada peringkat
- Fisiologia - Respiratoria - III TRANSPORTE DE GASESDokumen29 halamanFisiologia - Respiratoria - III TRANSPORTE DE GASEScristian duqueBelum ada peringkat
- INTERCAMBIO DE GASES EN EL PULMÓN Y TEJIDOSDokumen28 halamanINTERCAMBIO DE GASES EN EL PULMÓN Y TEJIDOSYesenia LopezBelum ada peringkat
- Presentacion Gratis Salud Medico Ilustracion CelesteDokumen25 halamanPresentacion Gratis Salud Medico Ilustracion CelesteJaclyn LozaniBelum ada peringkat
- Fisiología Respiratoria ResumenDokumen5 halamanFisiología Respiratoria Resumenarmando guitierrezBelum ada peringkat
- Capitulo 40Dokumen4 halamanCapitulo 40ArlenneRivera100% (2)
- Intercambio O2-CO2Dokumen5 halamanIntercambio O2-CO2Galo Pinos100% (1)
- Apunte4 ALTERACIONES DEL TRANSPORTE DE OXÍGENODokumen6 halamanApunte4 ALTERACIONES DEL TRANSPORTE DE OXÍGENODANIEL ALEJANDROBelum ada peringkat
- CAPITULO-40 Fisiologia de Guyton - ResumenDokumen4 halamanCAPITULO-40 Fisiologia de Guyton - ResumenMiguel Angel Manotas CañasBelum ada peringkat
- Introduccion Sistema Respiratorio PDFDokumen4 halamanIntroduccion Sistema Respiratorio PDFNicoleBelum ada peringkat
- FisicaDokumen5 halamanFisicaDenilsonCarlosAlaSamayaniBelum ada peringkat
- FISIOPATOLOGíA RESPIRATORIADokumen5 halamanFISIOPATOLOGíA RESPIRATORIAZELMA VALVERDE LAZAROBelum ada peringkat
- Transporte de Dioxido de Carbono y Oxigeno en La SangreDokumen30 halamanTransporte de Dioxido de Carbono y Oxigeno en La SangreEmilio Godoy MorenoBelum ada peringkat
- Respiratorio 3 Ventilacion PulmonarDokumen4 halamanRespiratorio 3 Ventilacion PulmonarFrancisco PabonBelum ada peringkat
- Transporte de Oxígeno y Dióxido de Carbono en La Sangre y Líquidos TisularesDokumen5 halamanTransporte de Oxígeno y Dióxido de Carbono en La Sangre y Líquidos TisularesDayanara RodriguezBelum ada peringkat
- Capitulo 1 La Respiración Conceptos y Alteraciones 1Dokumen9 halamanCapitulo 1 La Respiración Conceptos y Alteraciones 1Abigail CruzBelum ada peringkat
- El Sistema RespiratorioDokumen25 halamanEl Sistema RespiratorioCamila Alvarez MayorgaBelum ada peringkat
- Ventilación y PerfusiónDokumen16 halamanVentilación y Perfusióneli shagthBelum ada peringkat
- Perfusión 2021Dokumen17 halamanPerfusión 2021bdelkineBelum ada peringkat
- Relacion Ventilación - PerfusiónDokumen11 halamanRelacion Ventilación - PerfusiónJosmirys Rendon SifontesBelum ada peringkat
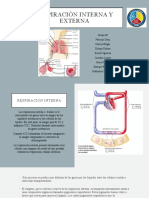
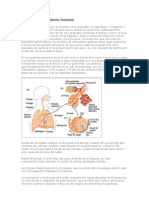
- Respiración Interna y ExternaDokumen9 halamanRespiración Interna y ExternaMia InestrozaBelum ada peringkat
- Transporte de Gases en La Sangre MORFO 2Dokumen37 halamanTransporte de Gases en La Sangre MORFO 2Ely CadenasBelum ada peringkat
- 8 Capitulo Portada 75Dokumen3 halaman8 Capitulo Portada 75Roberto Rouhana MonagasBelum ada peringkat
- Cap 41 - Transporte de O2 y Co2 en La Sangre y Liquidos TisularesDokumen9 halamanCap 41 - Transporte de O2 y Co2 en La Sangre y Liquidos TisularesJennifer RiveraBelum ada peringkat
- Oxigenoterapia FiiDokumen59 halamanOxigenoterapia FiiKarlos CórdovaBelum ada peringkat
- Relación Índice V/QDokumen24 halamanRelación Índice V/Qviconepg0% (2)
- El Sistema Respiratorio Humano-CurtisDokumen3 halamanEl Sistema Respiratorio Humano-CurtisAndrea Juarez100% (1)
- HematosisDokumen15 halamanHematosisTyet RbBelum ada peringkat
- Conceptos Y Estrategias En Terapia RespiratoriaDari EverandConceptos Y Estrategias En Terapia RespiratoriaPenilaian: 4 dari 5 bintang4/5 (1)
- Anatomia GeneralidadesDokumen161 halamanAnatomia GeneralidadesDailis Céspedes50% (2)
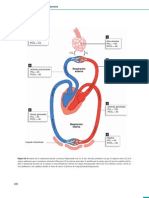
- Corazón y CirculaciónDokumen10 halamanCorazón y CirculaciónClaraFernandezBelum ada peringkat
- Adoum Lecciones Colegio de La Magos 2 ParteDokumen58 halamanAdoum Lecciones Colegio de La Magos 2 ParteYres Laqbar EdBelum ada peringkat
- Anatomía Parcial MacarenaDokumen63 halamanAnatomía Parcial MacarenaLucia Salvadora FernandezBelum ada peringkat
- ALBUMDokumen16 halamanALBUMMarly NiñoBelum ada peringkat
- Anatomía circulatoria y respiratoriaDokumen137 halamanAnatomía circulatoria y respiratoriaAna Sánchez AsensioBelum ada peringkat
- Circulacion Mayor y MenorDokumen3 halamanCirculacion Mayor y MenorNataly Cuevas100% (2)
- Manual de AnatomiaDokumen71 halamanManual de AnatomiaMiguel RoqueBelum ada peringkat
- Circulacion de Medula EspinalDokumen28 halamanCirculacion de Medula EspinalDamaris Peña KortingBelum ada peringkat
- Tromboembolismo Pulmonar: GenéticosDokumen5 halamanTromboembolismo Pulmonar: GenéticosMaria Camila RuizBelum ada peringkat
- Untitled 2Dokumen1 halamanUntitled 2María AvendañoBelum ada peringkat
- El CorazonDokumen2 halamanEl CorazonMabel RamirezBelum ada peringkat
- Sistema Cardiovascular Anatomia-1Dokumen85 halamanSistema Cardiovascular Anatomia-1MERY OBANDO100% (1)
- Exanguinotransfusión. APCDokumen6 halamanExanguinotransfusión. APCElena GomezBelum ada peringkat
- Funcionamiento corazónDokumen2 halamanFuncionamiento corazónewjorodi100% (1)
- RCIU: Restricción del crecimiento intrauterinoDokumen21 halamanRCIU: Restricción del crecimiento intrauterinoMisael Sanchez Valdivia100% (1)
- Aparato CirculatorioDokumen8 halamanAparato Circulatorioareli100% (1)
- Aparato cardiovascular generalidadesDokumen8 halamanAparato cardiovascular generalidadesyeilysaltBelum ada peringkat
- Termoregulacion y Adaptacion A La Vida ExtrauterinaDokumen30 halamanTermoregulacion y Adaptacion A La Vida ExtrauterinaJhamyle ZorelBelum ada peringkat
- Vias de Acceso Central y PerifericoDokumen21 halamanVias de Acceso Central y PerifericoMishelle Cercado RuizBelum ada peringkat
- El Sistema CardiovascularDokumen4 halamanEl Sistema CardiovascularCris PolancoBelum ada peringkat
- Pulso VenosoDokumen5 halamanPulso VenosoTatiana GomezBelum ada peringkat
- Evaluación Diagnóstica 2Dokumen2 halamanEvaluación Diagnóstica 2Edson Noel Flórez AlvaradoBelum ada peringkat
- Conceptos Básicos de RX de Tórax (Dra. Ruiz) PDFDokumen43 halamanConceptos Básicos de RX de Tórax (Dra. Ruiz) PDFYorman Arturo Gonzalez BallestasBelum ada peringkat
- Resumen Anatomía CorazónDokumen4 halamanResumen Anatomía CorazónVivian LoboBelum ada peringkat
- Sistema Venoso PortalDokumen4 halamanSistema Venoso PortalEric AvellanedaBelum ada peringkat
- Actividad Grupal..Dokumen4 halamanActividad Grupal..Aimee Villanueva CoronadoBelum ada peringkat
- Presion Venosa CentralDokumen8 halamanPresion Venosa CentralRenan Lopez ViañaBelum ada peringkat
- El Sistema CirculatorioDokumen9 halamanEl Sistema CirculatorioCarmen Chagua FabianBelum ada peringkat
- 1 Anatomia y Fisiologia Del CorazonDokumen5 halaman1 Anatomia y Fisiologia Del CorazonsaraiBelum ada peringkat