Anda mungkin juga menyukai
- Diplomado A Distancia de Medicina, Cirguia y Zootenia de Perros y GatosDokumen244 halamanDiplomado A Distancia de Medicina, Cirguia y Zootenia de Perros y GatosBrisbany AlcibarBelum ada peringkat
- TRIPTICO DiabetesDokumen2 halamanTRIPTICO DiabetesJorge Daniel Lopez Escobar50% (4)
- Cuestionario de NeuroanatomiaDokumen35 halamanCuestionario de NeuroanatomiaJonathan Montesdeoca97% (31)
- Precisiones dirigidas a la atención de estudiantes con variabilidades en el desarrolloDari EverandPrecisiones dirigidas a la atención de estudiantes con variabilidades en el desarrolloBelum ada peringkat
- SNAP-IV Abreviada PDFDokumen2 halamanSNAP-IV Abreviada PDFmanoilocBelum ada peringkat
- Guía Clínica para El Trastorno Negativista Desafiante - Josué Vasquéz, Miriam FeriaDokumen25 halamanGuía Clínica para El Trastorno Negativista Desafiante - Josué Vasquéz, Miriam FeriaIsrael Mendieta100% (5)
- Actualización Del Trastorno Por Déficit de Atención e Hiperactividad (Tdah) en NiñosDokumen48 halamanActualización Del Trastorno Por Déficit de Atención e Hiperactividad (Tdah) en NiñosMiguel ÁngelBelum ada peringkat
- Métodos de intervención Terapias más utilizadas en niños con autismoDari EverandMétodos de intervención Terapias más utilizadas en niños con autismoBelum ada peringkat
- Orientaciones Profesorado Altas CapacidadesDokumen7 halamanOrientaciones Profesorado Altas CapacidadesJose Luis MalagonBelum ada peringkat
- Transtorno Del Espectro Autista (4) .MAIRADokumen2 halamanTranstorno Del Espectro Autista (4) .MAIRAMay RMBelum ada peringkat
- Estrategias Pedagógicas para La DiversidadDokumen74 halamanEstrategias Pedagógicas para La DiversidadGeorgina52014Belum ada peringkat
- Entrevista NeuropsicológicaDokumen21 halamanEntrevista NeuropsicológicaADRIANABelum ada peringkat
- Neurodiversidad Discapacidad e Intelig MúltiplesDokumen6 halamanNeurodiversidad Discapacidad e Intelig MúltiplesJessica RiquelmeBelum ada peringkat
- 01MSEC ISenabre PDFDokumen69 halaman01MSEC ISenabre PDFNazaGutiérrezMartínezBelum ada peringkat
- 14 Rosselli - Desarrollo Habilidades Visoespaciales Enero Junio Vol 151 2015 PDFDokumen26 halaman14 Rosselli - Desarrollo Habilidades Visoespaciales Enero Junio Vol 151 2015 PDFPablo Alejandro Hermosilla RuizBelum ada peringkat
- Defensa TFM YADHIRA LEYTONDokumen33 halamanDefensa TFM YADHIRA LEYTONyadyBelum ada peringkat
- Juegos para Entrenar y Mejorar Dificultades en El TDAHDokumen4 halamanJuegos para Entrenar y Mejorar Dificultades en El TDAHIrene Cazorla TorresBelum ada peringkat
- Comorbilidad TDAH y AutismoDokumen2 halamanComorbilidad TDAH y AutismoJonathan GrandaBelum ada peringkat
- Funciones Ejecutivas y Su Importancia en Los Procesos de Atención, Memoria y AprendizajeDokumen9 halamanFunciones Ejecutivas y Su Importancia en Los Procesos de Atención, Memoria y AprendizajeAmy MartínezBelum ada peringkat
- Diapos Taller de MindfulnessDokumen12 halamanDiapos Taller de MindfulnesslilianaBelum ada peringkat
- WISC y Espectro AutistaDokumen150 halamanWISC y Espectro Autistaandrea ochoa100% (1)
- PuedoDokumen16 halamanPuedoMariel EstBelum ada peringkat
- TFM FinalDokumen30 halamanTFM FinalAndrea Gallo de la PazBelum ada peringkat
- Programa Sobre El Abordaje Al TDAHDokumen6 halamanPrograma Sobre El Abordaje Al TDAHAndrés Napo100% (1)
- Guia Estimulacion PDFDokumen14 halamanGuia Estimulacion PDFJuanyfeny Carmilema FajardoBelum ada peringkat
- La NeuroeducaciónDokumen4 halamanLa NeuroeducaciónMilagros Tapia0% (1)
- Modelo Informe Psic SÉNECADokumen7 halamanModelo Informe Psic SÉNECAnaimiguelBelum ada peringkat
- GuiaTDAH Profesores Modificada PDFDokumen42 halamanGuiaTDAH Profesores Modificada PDFCarolinaBelum ada peringkat
- Master Psicogia Sanitaria VIUDokumen2 halamanMaster Psicogia Sanitaria VIUpacojhidalgo0% (1)
- AEmociones y AutismoDokumen8 halamanAEmociones y AutismoPaulina Andrea Núñez Soto100% (1)
- PDF Tea Intervención Sep.Dokumen11 halamanPDF Tea Intervención Sep.Fátima PérezBelum ada peringkat
- Estrategias para Evitar El Deterioro Cognitivo en AdultosDokumen9 halamanEstrategias para Evitar El Deterioro Cognitivo en AdultosKeyla GarciaBelum ada peringkat
- AcaiDokumen53 halamanAcaiSabrina SaloBelum ada peringkat
- Rehabilitacion NeuropsicologicaDokumen9 halamanRehabilitacion NeuropsicologicaCarlos MilhasBelum ada peringkat
- Trastorno de AspergerDokumen14 halamanTrastorno de AspergerDany Sf TabernaBelum ada peringkat
- NeuromitosDokumen6 halamanNeuromitosPauli Sáez KortmannBelum ada peringkat
- Dificultades de Aprendizaje - Organized PDFDokumen53 halamanDificultades de Aprendizaje - Organized PDFJoaquin FigueroaBelum ada peringkat
- Trabajo Practico HiperactividadDokumen30 halamanTrabajo Practico HiperactividadDANIEL ALEJANDRO100% (1)
- 2017 - Economía de Fichas en TEADokumen53 halaman2017 - Economía de Fichas en TEASilvia100% (1)
- Discapacidad IntelectualDokumen13 halamanDiscapacidad IntelectualDamaguiBelum ada peringkat
- TDAH Recopilacion de DatosDokumen54 halamanTDAH Recopilacion de DatosAlan BlondBelum ada peringkat
- Guia de Intervencion Educativa en Trastornos de ConductaDokumen30 halamanGuia de Intervencion Educativa en Trastornos de ConductajuanBelum ada peringkat
- TFG PrietoGarcia DepresionPostpartoDokumen33 halamanTFG PrietoGarcia DepresionPostpartoivanna villavicencioBelum ada peringkat
- Unidad 1 Rol Del Neuropsicologo Infantil y Evaluación NeuropsicológicaDokumen29 halamanUnidad 1 Rol Del Neuropsicologo Infantil y Evaluación Neuropsicológicacarola Tórrez MéridaBelum ada peringkat
- Angel Daniel Reporte de Evaluación NeuropsicológicaDokumen3 halamanAngel Daniel Reporte de Evaluación NeuropsicológicaAngel Daniel VásquezBelum ada peringkat
- Teoría de La Mente en Niños de Corta EdadDokumen3 halamanTeoría de La Mente en Niños de Corta EdadCamila PazBelum ada peringkat
- Apoyo Conductual Positivo - Manuales de Trabajo en Centros de Atención A Personas Con Discapacidad de La Junta de Castilla y LeónDokumen55 halamanApoyo Conductual Positivo - Manuales de Trabajo en Centros de Atención A Personas Con Discapacidad de La Junta de Castilla y LeónMónica LorenzoBelum ada peringkat
- Psicología y Ceguera 003Dokumen220 halamanPsicología y Ceguera 003Jorge Luis Fernandez SalgueroBelum ada peringkat
- Girls and Women With Austism - En.esDokumen16 halamanGirls and Women With Austism - En.esCarito Hernandez RodriguezBelum ada peringkat
- Expo Sindrome DisejecutivoDokumen9 halamanExpo Sindrome DisejecutivoPaola Andrea RODRIGUEZ QUEVEDOBelum ada peringkat
- Tema 4 Desarrollo de Las Competencias Cognitivas (Modo de Compatibilidad)Dokumen26 halamanTema 4 Desarrollo de Las Competencias Cognitivas (Modo de Compatibilidad)dolores ecijaBelum ada peringkat
- Manual-Herramientas Inclusion. ComplementaDokumen69 halamanManual-Herramientas Inclusion. ComplementaNatalia HerreraBelum ada peringkat
- Trastornos Mentales Organicos IIDokumen48 halamanTrastornos Mentales Organicos IIapi-3705495100% (3)
- Tema 2 - Evaluación de Los Trastornos Del NeurodesarrolloDokumen9 halamanTema 2 - Evaluación de Los Trastornos Del Neurodesarrollomarfil123Belum ada peringkat
- Antologia Atencion A Las DiscapacidadesDokumen102 halamanAntologia Atencion A Las Discapacidadesfrx22Belum ada peringkat
- Análisis de Las Fases en La Enfermedad de AlzheimerDokumen13 halamanAnálisis de Las Fases en La Enfermedad de AlzheimeryadyBelum ada peringkat
- Desarrollo de Habilidades PsicosocialesDokumen3 halamanDesarrollo de Habilidades PsicosocialesSantii VilchesBelum ada peringkat
- Evaluación de La Eficacia de Las Intervenciones Psicoeducativas en Los Trastornos Del Espectro AutistaDokumen142 halamanEvaluación de La Eficacia de Las Intervenciones Psicoeducativas en Los Trastornos Del Espectro AutistaEquipo de Orientación Liceo CastillaBelum ada peringkat
- El mediador comunicativo en el equipo interdisciplinar educativo. SSCG0211Dari EverandEl mediador comunicativo en el equipo interdisciplinar educativo. SSCG0211Belum ada peringkat
- Intervención con familias de personas con discapacidad. SSCE0111Dari EverandIntervención con familias de personas con discapacidad. SSCE0111Belum ada peringkat
- Buenas prácticas inclusivas en la respuesta al alumnado con enfermedades raras: Estudio en diferentes conunidads autónomasDari EverandBuenas prácticas inclusivas en la respuesta al alumnado con enfermedades raras: Estudio en diferentes conunidads autónomasBelum ada peringkat
- Manual Metodologico para el Pedagogo EspecialDari EverandManual Metodologico para el Pedagogo EspecialBelum ada peringkat
- Aplicación de los programas de habilidades de autonomía personal y social del ACNEE. SSCE0112Dari EverandAplicación de los programas de habilidades de autonomía personal y social del ACNEE. SSCE0112Belum ada peringkat
- Perfil de las Inteligencias Múltiples de los Niños del Nivel Inicial 2Dari EverandPerfil de las Inteligencias Múltiples de los Niños del Nivel Inicial 2Belum ada peringkat
- Estructura Del Sistema NerviosoDokumen81 halamanEstructura Del Sistema NerviosoFernando Gil50% (2)
- Diagnóstico y Tratamiento Del Trastorno Por Déficit de Atención-Hiperactividad en Adultos PDFDokumen11 halamanDiagnóstico y Tratamiento Del Trastorno Por Déficit de Atención-Hiperactividad en Adultos PDFmax_reyeslobos100% (1)
- PIENSA Guia Adolescentes Psicosis PDFDokumen142 halamanPIENSA Guia Adolescentes Psicosis PDFJordi Cañadas LopezBelum ada peringkat
- Tecnicas para Cartas de Amor U OdioDokumen4 halamanTecnicas para Cartas de Amor U OdioChristian Gabriel Stampacchio100% (16)
- EsquizofreniaDokumen9 halamanEsquizofreniamanoilocBelum ada peringkat
- Art TdahDokumen7 halamanArt TdahMargarita ValenciaBelum ada peringkat
- Estructura Del Sistema NerviosoDokumen81 halamanEstructura Del Sistema NerviosoFernando Gil50% (2)
- Psicopat Del PensmaientoDokumen13 halamanPsicopat Del PensmaientovasuzuBelum ada peringkat
- Curso Terapia Basada en La Mentalización PDFDokumen25 halamanCurso Terapia Basada en La Mentalización PDFcerezaroja3386100% (4)
- Psicopat Del PensmaientoDokumen13 halamanPsicopat Del PensmaientovasuzuBelum ada peringkat
- Analisis de Redes en Psicología PDFDokumen12 halamanAnalisis de Redes en Psicología PDFmanoilocBelum ada peringkat
- H.2 TICS Spanish 2018Dokumen15 halamanH.2 TICS Spanish 2018Ptrc Lbr Lp100% (1)
- Guia PanicoDokumen16 halamanGuia PanicoVanessa Gallardo YáñezBelum ada peringkat
- PIENSA Guia Adolescentes Psicosis PDFDokumen142 halamanPIENSA Guia Adolescentes Psicosis PDFJordi Cañadas LopezBelum ada peringkat
- Pautas Manejo Disrupción AulaDokumen2 halamanPautas Manejo Disrupción AulamanoilocBelum ada peringkat
- Pródomos Psicosis AdolescenteDokumen14 halamanPródomos Psicosis AdolescentemanoilocBelum ada peringkat
- ¿Quién Te Quiere A Tí?Dokumen60 halaman¿Quién Te Quiere A Tí?Colegio de Alpartir100% (1)
- Tecnicas para Cartas de Amor U OdioDokumen4 halamanTecnicas para Cartas de Amor U OdioChristian Gabriel Stampacchio100% (16)
- Trastorno Bipolar en La Infancia y AdolescenciaDokumen13 halamanTrastorno Bipolar en La Infancia y AdolescenciatedansaBelum ada peringkat
- Tecnicas para Cartas de Amor U OdioDokumen4 halamanTecnicas para Cartas de Amor U OdioChristian Gabriel Stampacchio100% (16)
- Terapia de Esquemas PDFDokumen16 halamanTerapia de Esquemas PDFLuis Enrique Ferrúa Garcia100% (1)
- Guía Infanciaadolescente PDFDokumen84 halamanGuía Infanciaadolescente PDFDorita CalelBelum ada peringkat
- Guía Infanciaadolescente PDFDokumen84 halamanGuía Infanciaadolescente PDFDorita CalelBelum ada peringkat
- Tecnicas para Cartas de Amor U OdioDokumen4 halamanTecnicas para Cartas de Amor U OdioChristian Gabriel Stampacchio100% (16)
- Los Trastornos de Personalidad Según Millon (2007)Dokumen1 halamanLos Trastornos de Personalidad Según Millon (2007)manoilocBelum ada peringkat
- Información Sobre El PerroDokumen19 halamanInformación Sobre El Perrovanessardz1Belum ada peringkat
- Ataques Al Vínculo - W. J. BionDokumen18 halamanAtaques Al Vínculo - W. J. Bioneduardojev100% (1)
- Ficha de Investigación Epidemiológica de Caso de Sífilis Materna y Sífilis CongénitaDokumen2 halamanFicha de Investigación Epidemiológica de Caso de Sífilis Materna y Sífilis CongénitaNadia Sánchez LizanaBelum ada peringkat
- Resumen Historia de La Medicina - by Alan A. Garcia Desperés y Luigi GarcíaDokumen57 halamanResumen Historia de La Medicina - by Alan A. Garcia Desperés y Luigi GarcíaLara Agostina GabarruBelum ada peringkat
- Ficha Tecnica PrexorDokumen1 halamanFicha Tecnica Prexorhuguini007Belum ada peringkat
- SECUENCIA 1 Características de Los Seres VivosDokumen22 halamanSECUENCIA 1 Características de Los Seres VivosMara LoplanBelum ada peringkat
- CampylobacterDokumen33 halamanCampylobacterAdelheint Bernstein100% (1)
- GastritisDokumen8 halamanGastritisMariano Duarte EcheverriaBelum ada peringkat
- OFIDIOSDokumen6 halamanOFIDIOSRamonaNoviembreBelum ada peringkat
- Estrategia de TBCCCCDokumen108 halamanEstrategia de TBCCCCSxmpk FV0% (1)
- Presentacion - Factores de Riesgo de PeriimplantitisDokumen17 halamanPresentacion - Factores de Riesgo de PeriimplantitisFredy'n De RodríguezBelum ada peringkat
- Pericarditis y TaponamientoDokumen53 halamanPericarditis y TaponamientojurgenBelum ada peringkat
- Ficha Farmacologica Nombre Generico - ComDokumen8 halamanFicha Farmacologica Nombre Generico - ComJal Jeyson Alania RiosBelum ada peringkat
- CPO EpidemiologíaDokumen5 halamanCPO EpidemiologíaFannyBelum ada peringkat
- Manejo Odontológico Del Paciente EpilepticoDokumen20 halamanManejo Odontológico Del Paciente EpilepticoAlejandra Guallo PerezBelum ada peringkat
- Examen Físico de AbdomenDokumen5 halamanExamen Físico de AbdomenMarcelysBelum ada peringkat
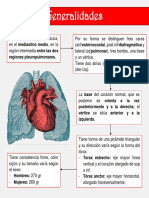
- CorazónDokumen4 halamanCorazónPaula AlvaradoBelum ada peringkat
- Syllabus FinalDokumen26 halamanSyllabus FinalChristofer George BarreraBelum ada peringkat
- Revista Bellatriz 6ta EdiciónDokumen24 halamanRevista Bellatriz 6ta EdiciónservisualBelum ada peringkat
- NK Feb 23 PDFDokumen27 halamanNK Feb 23 PDFLaura Sofia Sorza MarinBelum ada peringkat
- Modelo de Informe de Auditoria de CasoDokumen6 halamanModelo de Informe de Auditoria de CasoIRINA FERNANDEZ PISFILBelum ada peringkat
- FuatDokumen1 halamanFuatGRECIABelum ada peringkat
- Enfermedad Celiaca: Revisión: Celiac Disease. A ReviewDokumen15 halamanEnfermedad Celiaca: Revisión: Celiac Disease. A ReviewErick CocoletziBelum ada peringkat
- AntibioticosDokumen25 halamanAntibioticosPasoloco1Belum ada peringkat
- Guia de Valoración Por Dominio ContiDokumen8 halamanGuia de Valoración Por Dominio ContiRosario Del Pilar Papuico ManriqueBelum ada peringkat
- Cuidados Inmediatos y Mediatos Al Recién NacidoDokumen14 halamanCuidados Inmediatos y Mediatos Al Recién NacidoDiego Lopez PiñaBelum ada peringkat
- Manual de Cocina Saludable Trimestral (Recuperado)Dokumen72 halamanManual de Cocina Saludable Trimestral (Recuperado)Dení BuendíaBelum ada peringkat