Anda mungkin juga menyukai
- 10-Bioquimica 27 Octubre FacultadDokumen31 halaman10-Bioquimica 27 Octubre FacultadVizcaino Rojas JosèBelum ada peringkat
- Proyecto de InvestigacionDokumen19 halamanProyecto de InvestigacionPercy Perez50% (2)
- Beta OxidaciónDokumen12 halamanBeta OxidaciónIvi100% (1)
- Bioquimica Proteinas Plasmaticas TerminadoDokumen15 halamanBioquimica Proteinas Plasmaticas Terminadoerick marioBelum ada peringkat
- GlucogenolisisDokumen47 halamanGlucogenolisisDannerPlay25 20Belum ada peringkat
- Sintesis de Acidos GrasosDokumen41 halamanSintesis de Acidos GrasosDiego Alejandro Cuevas Lau100% (1)
- Qué Son Los Aminoácidos EsencialesDokumen2 halamanQué Son Los Aminoácidos EsencialesMaría ChavezBelum ada peringkat
- Síntesis de Ácidos GrasosDokumen17 halamanSíntesis de Ácidos GrasosBelén Chamorro SevillaBelum ada peringkat
- Tema 3 Metabolismo Lipidos 111 2 ParcialDokumen30 halamanTema 3 Metabolismo Lipidos 111 2 ParcialMario BownstherBelum ada peringkat
- Lipolisis y B-OxidaciónDokumen34 halamanLipolisis y B-OxidaciónJORGE ANTONIO GONZALEZ BRAVO100% (1)
- Salud mental de hombres mayores en Chile: una realidad por priorizarDokumen5 halamanSalud mental de hombres mayores en Chile: una realidad por priorizarDaniela Campos HerreraBelum ada peringkat
- Evaluación neuropsicológica infantil con el CUMANINDokumen22 halamanEvaluación neuropsicológica infantil con el CUMANINPsicología 2do año100% (2)
- Rutas MetabolicasDokumen30 halamanRutas Metabolicasamilcar torresBelum ada peringkat
- Metabolismo de Los Aminoácidos y de Las ProteínasDokumen18 halamanMetabolismo de Los Aminoácidos y de Las ProteínasAnthony Diaz0% (1)
- 23) Resumen (Caballo) Capítulo 24 - Terapia Cognitivo Estructural, El Modelo de Guidano y LiottiDokumen2 halaman23) Resumen (Caballo) Capítulo 24 - Terapia Cognitivo Estructural, El Modelo de Guidano y LiottiMariajesus Hernandez SalinasBelum ada peringkat
- Tema13 Catabolismo AminoacidosDokumen36 halamanTema13 Catabolismo AminoacidosÁngela Díaz ChamorroBelum ada peringkat
- Digestión, Absorción y Metabolismo de Los CarbohidratosDokumen51 halamanDigestión, Absorción y Metabolismo de Los CarbohidratosMeri Y Sami GómezBelum ada peringkat
- Ensayo Acidos GrasosDokumen3 halamanEnsayo Acidos GrasosKro BrtlBelum ada peringkat
- Aminoacidos No Esenciales (Expo)Dokumen16 halamanAminoacidos No Esenciales (Expo)AlbaOsorioBelum ada peringkat
- w4 Clase Bioca Cetogenesis Cetolisis Xenob 20052020Dokumen32 halamanw4 Clase Bioca Cetogenesis Cetolisis Xenob 20052020Darreem Steve Villarreal PutnamBelum ada peringkat
- LIPOLISISDokumen8 halamanLIPOLISISScarlette OlivoBelum ada peringkat
- Beta OxidaciónDokumen18 halamanBeta OxidaciónBryan SantanaBelum ada peringkat
- Compuestos NitrogenadosDokumen14 halamanCompuestos NitrogenadosCaroline TW0% (1)
- TEMA 46 Biosintesis de Ácidos GrasosDokumen6 halamanTEMA 46 Biosintesis de Ácidos GrasosAkimita100% (3)
- Catabolismo de AminoacidosDokumen20 halamanCatabolismo de AminoacidosXimena Véliz DaudBelum ada peringkat
- Ciclo Del Ácido TricarboxílicoDokumen32 halamanCiclo Del Ácido TricarboxílicoDenissLopez100% (1)
- Fosforilación OxidativaDokumen4 halamanFosforilación OxidativaDaleska C. GalarzaBelum ada peringkat
- El ciclo de la urea: transformación del amoníaco en ureaDokumen65 halamanEl ciclo de la urea: transformación del amoníaco en ureaAlexandraMedinaCastilloBelum ada peringkat
- Guia LipoproteinasDokumen16 halamanGuia LipoproteinasJesu Herrera67% (3)
- Metabolismo de carbohidratos y lípidosDokumen4 halamanMetabolismo de carbohidratos y lípidosDaniela RoseroBelum ada peringkat
- Ciclo de Krebs resumen: las 8 reacciones claveDokumen1 halamanCiclo de Krebs resumen: las 8 reacciones claveMaría Trinidad Fraga YoliBelum ada peringkat
- Ciclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoDokumen7 halamanCiclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoJeanette PaulinaBelum ada peringkat
- Glucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoDokumen26 halamanGlucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoMarcelo Kim100% (1)
- Bioquimica de La Diabetes 2Dokumen16 halamanBioquimica de La Diabetes 2AdonayDecoraciones50% (2)
- Unidad 3 Metabolismo de NutrientesDokumen6 halamanUnidad 3 Metabolismo de NutrientesfernandoalvarezdelccBelum ada peringkat
- Rutas metabólicasDokumen2 halamanRutas metabólicasAlvaro Hernández100% (1)
- Glucólisis: La ruta universal para extraer energía de la glucosaDokumen21 halamanGlucólisis: La ruta universal para extraer energía de la glucosatatahimys0% (1)
- GlucogenólisisDokumen3 halamanGlucogenólisisJosué Delgado Navas100% (1)
- METABOLISMO GLUCOGENODokumen34 halamanMETABOLISMO GLUCOGENOEsmeralda CarreraBelum ada peringkat
- Sistema CardiovascularDokumen12 halamanSistema CardiovascularCristina VelázquezBelum ada peringkat
- Los carbohidratos: fuente de energía y componentes estructuralesDokumen4 halamanLos carbohidratos: fuente de energía y componentes estructuralesNicole AndradeBelum ada peringkat
- Guía de Bioquímica. LípidosDokumen27 halamanGuía de Bioquímica. LípidosAlfonso RodríguezBelum ada peringkat
- Metabolismo Microbiano Ruminal y Enfermedades MetabólicasDokumen18 halamanMetabolismo Microbiano Ruminal y Enfermedades MetabólicasvalentinaBelum ada peringkat
- Cuestionario 11Dokumen2 halamanCuestionario 11Mayker PanibraBelum ada peringkat
- Estructura y modificaciones del almidónDokumen6 halamanEstructura y modificaciones del almidónHeber Cordero EguiaBelum ada peringkat
- Ciclo de KrebsDokumen6 halamanCiclo de KrebsAguila PaulBelum ada peringkat
- Cap. 18-Metabolismo de NucleótidosDokumen9 halamanCap. 18-Metabolismo de NucleótidosNatalia PinedaBelum ada peringkat
- Ciclo de La UreaDokumen5 halamanCiclo de La UreaMartinezHuertaMaryBelum ada peringkat
- ENZIMASDokumen38 halamanENZIMASClaudia LoredoBelum ada peringkat
- Glucolisis 21645 PDFDokumen77 halamanGlucolisis 21645 PDFShärlÿ ContrerasBelum ada peringkat
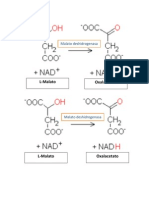
- La Enzima Malato DeshidrogenasaDokumen2 halamanLa Enzima Malato DeshidrogenasaKaren Torres JuárezBelum ada peringkat
- Ciclo de la urea: reacciones y regulaciónDokumen3 halamanCiclo de la urea: reacciones y regulaciónlelsiBelum ada peringkat
- 15 Digestión, Absorción y Transporte de Lípidos Por Las LipoproteínasDokumen8 halaman15 Digestión, Absorción y Transporte de Lípidos Por Las LipoproteínasMacarena Fernanda LizamaBelum ada peringkat
- Isiologia Del Metabolismo de La Proteinas PDFDokumen19 halamanIsiologia Del Metabolismo de La Proteinas PDFRosario Laguna100% (1)
- Oxidacion de Ácidos GrasosDokumen1 halamanOxidacion de Ácidos Grasosagustin zarate cruzBelum ada peringkat
- Vía de Las PentosasDokumen30 halamanVía de Las PentosasAlexia Fuentes100% (1)
- Biosíntesis de Carbohidratos - BIOQUIMICA II U2Dokumen13 halamanBiosíntesis de Carbohidratos - BIOQUIMICA II U2Bernardo Morales0% (1)
- Óxido Nítrico SintasaDokumen4 halamanÓxido Nítrico SintasaKarina ParkBelum ada peringkat
- CETOGÉNESISDokumen15 halamanCETOGÉNESISINick Salva PastorBelum ada peringkat
- Cuadros LipolisisDokumen8 halamanCuadros LipolisisLFNA SoporteBelum ada peringkat
- Biosíntesis de ColesterolDokumen6 halamanBiosíntesis de ColesterolAlejandra ViteriBelum ada peringkat
- El ciclo de Krebs, ruta central del metabolismo aerobioDokumen48 halamanEl ciclo de Krebs, ruta central del metabolismo aerobioVerónica LoaizaBelum ada peringkat
- Fisiopatologia Anemia MegaloblasticaDokumen5 halamanFisiopatologia Anemia MegaloblasticaAlex MirandaBelum ada peringkat
- Síntesis de Ácidos GrasosDokumen19 halamanSíntesis de Ácidos GrasosOriana RiveroBelum ada peringkat
- Todos Los Documentos Del Caso Margarita CcahuanaDokumen35 halamanTodos Los Documentos Del Caso Margarita CcahuanaFra Johanna GallegosBelum ada peringkat
- BOLETIN I SEMESTRE - 2023-OficialDokumen53 halamanBOLETIN I SEMESTRE - 2023-OficialNicol Medina MedinaBelum ada peringkat
- SPSU-855 Entregable02Dokumen4 halamanSPSU-855 Entregable02Cristian Nuñez BecerraBelum ada peringkat
- Infografía Durmiendo Con El Enemigo - Kassandra FedaltoDokumen1 halamanInfografía Durmiendo Con El Enemigo - Kassandra FedaltoalumniupcBelum ada peringkat
- Escalas de Contenido de Mmpi - 2Dokumen6 halamanEscalas de Contenido de Mmpi - 2Ingrid AZBelum ada peringkat
- Síntesis de Glucógeno J Grasas y Proteínas J Metabolismo J Calor y Vitaminas HidrosolublesDokumen12 halamanSíntesis de Glucógeno J Grasas y Proteínas J Metabolismo J Calor y Vitaminas HidrosolublesHaide BlancoBelum ada peringkat
- Enloqueciendo A KurtDokumen536 halamanEnloqueciendo A KurtIxia HidalgoBelum ada peringkat
- Taller Unidad 1Dokumen3 halamanTaller Unidad 1Ana Maria HernandezBelum ada peringkat
- Gestión Seguridad Salud Trabajo ConstrucciónDokumen18 halamanGestión Seguridad Salud Trabajo ConstrucciónJULIO ALDHAIR URIBE MORRISBelum ada peringkat
- 059 - Tratamiento Quirúrgico de Las Sinusitis y de Sus Complicaciones Cirugía de Abordaje Externo. Cirugía deDokumen24 halaman059 - Tratamiento Quirúrgico de Las Sinusitis y de Sus Complicaciones Cirugía de Abordaje Externo. Cirugía deFernando RiveraBelum ada peringkat
- Métodos de Planificación FamiliarDokumen10 halamanMétodos de Planificación FamiliarAngie LeodegarioBelum ada peringkat
- Sitio Arqueológico Mixco ViejoDokumen7 halamanSitio Arqueológico Mixco ViejoDylou Morales GámezBelum ada peringkat
- Guia Tne en EspañolDokumen33 halamanGuia Tne en EspañolmonicaBelum ada peringkat
- Mejoramiento del sistema de agua y desagüe en Pariamarca-CantaDokumen23 halamanMejoramiento del sistema de agua y desagüe en Pariamarca-CantaEnrique Gerardo Arbaiza ChavezBelum ada peringkat
- 5 Guayacan de ManizalesDokumen1 halaman5 Guayacan de ManizalesMaria Fernanda Riano PelaezBelum ada peringkat
- ?6° S12 ACTIVIDADES SEMANALES Esmeralda Te EnseñaDokumen20 halaman?6° S12 ACTIVIDADES SEMANALES Esmeralda Te EnseñaYandil Gomez100% (1)
- Treck Curso Teorico y Practico Altura Física 08 HorasDokumen15 halamanTreck Curso Teorico y Practico Altura Física 08 Horasfrancisco alfaroBelum ada peringkat
- Formato de Declaracion de Accidentes Personales EstudiantesDokumen2 halamanFormato de Declaracion de Accidentes Personales EstudiantesPaul Ortiz PimentelBelum ada peringkat
- ENSAYO Lectoescritura SenaDokumen7 halamanENSAYO Lectoescritura SenaMARIA VALENTINA ARANA VASQUEZBelum ada peringkat
- ESCOLIOSISDokumen12 halamanESCOLIOSISAnnelys SotilloBelum ada peringkat
- Caso Clinico 3Dokumen13 halamanCaso Clinico 3Evelyn Sanchez MoreyraBelum ada peringkat
- Perfil TemperamentalDokumen4 halamanPerfil TemperamentalMyrelle FelicianoBelum ada peringkat
- Punciones Por MicrometodoDokumen3 halamanPunciones Por MicrometodovalentinaBelum ada peringkat
- Examen - (APEB1-15%) Resolución de Caso 1 - Responda El Cuestionario Planteado en Base Al Análisis de La Película "TOC TOC"Dokumen5 halamanExamen - (APEB1-15%) Resolución de Caso 1 - Responda El Cuestionario Planteado en Base Al Análisis de La Película "TOC TOC"Wladimir Coloma CruzBelum ada peringkat
- Enfoques de Atención Integral de SaludDokumen5 halamanEnfoques de Atención Integral de SaludRoss M RVBelum ada peringkat
- Tarea III Parctica de Psicoligia Clinica 2Dokumen13 halamanTarea III Parctica de Psicoligia Clinica 2mirquella ortizBelum ada peringkat