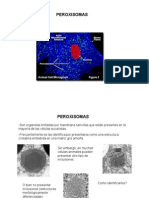
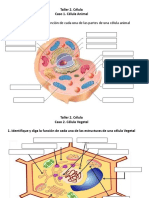
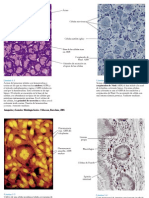
Anda mungkin juga menyukai
- Ciclo CelularDokumen37 halamanCiclo CelularAurelia Gonzales Calle100% (2)
- Biologia Practica 02 OBSERVACIÓN DE PREPARADOS EN FRECO Y EN SECODokumen6 halamanBiologia Practica 02 OBSERVACIÓN DE PREPARADOS EN FRECO Y EN SECONelson Aranibar100% (1)
- TranscripciónDokumen28 halamanTranscripciónAurelia Gonzales Calle100% (2)
- TranscripciónDokumen28 halamanTranscripciónAurelia Gonzales Calle100% (2)
- Fisiologia CelularDokumen57 halamanFisiologia CelularSoledad Loayza PumaBelum ada peringkat
- Fisiología Osteomioarticular Huesos, Musculos y ArticulacionesDokumen39 halamanFisiología Osteomioarticular Huesos, Musculos y ArticulacionesNicole Alexandra Garavito Nu�ezBelum ada peringkat
- La CélulaDokumen24 halamanLa CélulaIsabel Lermo BenitesBelum ada peringkat
- Taller Celula ValidacionDokumen4 halamanTaller Celula Validacionjhon fredy urbano lozanoBelum ada peringkat
- Guía de Estudio #4 Reticulo EndoplasmicoDokumen2 halamanGuía de Estudio #4 Reticulo EndoplasmicoW IngenieríaBelum ada peringkat
- Cadena Transportadora de ElectronesDokumen9 halamanCadena Transportadora de ElectronesVinicio VillafuerteBelum ada peringkat
- Aparato de GolgiDokumen9 halamanAparato de GolgiBrian RodeloBelum ada peringkat
- MetabolismoDokumen27 halamanMetabolismoLibros Andrea100% (2)
- Aparato de Golgi: estructura, funciones y transporte de vesículasDokumen6 halamanAparato de Golgi: estructura, funciones y transporte de vesículasSealim ChileBelum ada peringkat
- Carbohidratos UPAO ChunaDokumen30 halamanCarbohidratos UPAO ChunaIair AlessandroBelum ada peringkat
- Tema ProteinasDokumen28 halamanTema ProteinasAurelia Gonzales CalleBelum ada peringkat
- Tema ProteinasDokumen28 halamanTema ProteinasAurelia Gonzales CalleBelum ada peringkat
- Aparato de Golgi Lisosomas y VacuolasDokumen43 halamanAparato de Golgi Lisosomas y VacuolasscribdenesimoBelum ada peringkat
- RibosomasDokumen8 halamanRibosomasAurelia Gonzales Calle100% (1)
- Clase 6 - Transporte de MembranaDokumen24 halamanClase 6 - Transporte de MembranaDianita MendozaBelum ada peringkat
- Enzimas: Características y ClasificaciónDokumen27 halamanEnzimas: Características y ClasificaciónvictorBelum ada peringkat
- Ribosomas, RE, AGDokumen29 halamanRibosomas, RE, AGCarol FloresBelum ada peringkat
- Trabajo 3 de BiologiaDokumen22 halamanTrabajo 3 de BiologiaLeidy Tantaleán100% (1)
- Meiosis - GametogenesisDokumen32 halamanMeiosis - GametogenesisDianita MendozaBelum ada peringkat
- ReplicacionDokumen28 halamanReplicacionAurelia Gonzales CalleBelum ada peringkat
- Endosomas y LisosomasDokumen24 halamanEndosomas y LisosomasAurelia Gonzales Calle93% (14)
- PeroxisomasDokumen8 halamanPeroxisomasAurelia Gonzales CalleBelum ada peringkat
- Mediciones e IncertidumbreDokumen6 halamanMediciones e IncertidumbreSOREY SOLANGE PANDURO MARINABelum ada peringkat
- Compuestos OxigenadosDokumen11 halamanCompuestos OxigenadosemiliaGS2Belum ada peringkat
- Informe Re GolgiDokumen7 halamanInforme Re GolgienriqueBelum ada peringkat
- Reticulo Endoplasmatico LisoDokumen6 halamanReticulo Endoplasmatico LisoAndreaBelum ada peringkat
- 4-6 Difusión y Transporte Pasivo PDFDokumen5 halaman4-6 Difusión y Transporte Pasivo PDFBrayan Andrės CerónBelum ada peringkat
- Preguntas Resueltos - Los GlucidosDokumen5 halamanPreguntas Resueltos - Los Glucidosandrea riuBelum ada peringkat
- Examen ParcialDokumen3 halamanExamen ParcialKEVIN ENDER TAYPE HUANCABelum ada peringkat
- Seminario MutacionesDokumen46 halamanSeminario MutacionesDavid SiguenzaBelum ada peringkat
- Ácidos carboxílicos y sus derivadosDokumen44 halamanÁcidos carboxílicos y sus derivadosErikaNC796Belum ada peringkat
- PeroxisomasDokumen20 halamanPeroxisomasThuBhebesithaEmelexistaBelum ada peringkat
- Transporte de proteínas en la célulaDokumen4 halamanTransporte de proteínas en la célulaEsteban RuedaBelum ada peringkat
- T-37 Traducción Sintesis de ProteinasDokumen24 halamanT-37 Traducción Sintesis de ProteinasAnonymous iIaMAfVBelum ada peringkat
- Todo Polipeptido Es Una Proteina BiologíaDokumen1 halamanTodo Polipeptido Es Una Proteina BiologíaAndre Arcondo Lopez100% (1)
- Actividad de Reforzamiento y Nivelación 1 1Dokumen8 halamanActividad de Reforzamiento y Nivelación 1 1Fernando RodriguezBelum ada peringkat
- Tsa2 Proteinas - NRC 8970 - Diana&valeriDokumen8 halamanTsa2 Proteinas - NRC 8970 - Diana&valeriValeria Sandoval BancesBelum ada peringkat
- Ejercicios MEIOSIS Y GAMETOGÉNESIS PDFDokumen5 halamanEjercicios MEIOSIS Y GAMETOGÉNESIS PDFAbraham RamirezBelum ada peringkat
- Organelas 20citoplasmaticas 2013Dokumen17 halamanOrganelas 20citoplasmaticas 2013Luilly FernandoBelum ada peringkat
- Sistema de EndomembranasDokumen27 halamanSistema de EndomembranasSofía TorresBelum ada peringkat
- Traduccion Del ArnDokumen2 halamanTraduccion Del ArnDiana CampoverdeBelum ada peringkat
- PeroxisomasDokumen6 halamanPeroxisomasAndreaBelum ada peringkat
- s5 - PPT. 5.2. - Metabolismo de Las PorfirinasDokumen50 halamans5 - PPT. 5.2. - Metabolismo de Las PorfirinasCielo Chavez BarreraBelum ada peringkat
- Observacion de Organelos y Cromosomas CitoplasmaticosDokumen26 halamanObservacion de Organelos y Cromosomas CitoplasmaticosAlonso Paucar Diaz50% (2)
- TrombocitopeniaDokumen86 halamanTrombocitopeniaiberzBelum ada peringkat
- Formas de núcleo en células sanguíneasDokumen4 halamanFormas de núcleo en células sanguíneasDaniel Marquiño Alonzo Paz50% (2)
- R1-04-F-15-02-Transporte - A - Traves - de - La - Membrana - Celular PDFDokumen6 halamanR1-04-F-15-02-Transporte - A - Traves - de - La - Membrana - Celular PDFCristian CarpioBelum ada peringkat
- ADN Mitocondrial en Medicina ForenceDokumen9 halamanADN Mitocondrial en Medicina Forenceapi-3766024100% (1)
- Transcripcion y Traduccion de AdnDokumen17 halamanTranscripcion y Traduccion de AdnPercy Ureta SierraBelum ada peringkat
- NucleoDokumen20 halamanNucleoAurelia Gonzales CalleBelum ada peringkat
- Organelos CelularesDokumen9 halamanOrganelos CelularesPaula RodriguezBelum ada peringkat
- 20170908220928 (1)Dokumen41 halaman20170908220928 (1)Evelin Ramos MarcelianoBelum ada peringkat
- Necrosis I 2015Dokumen86 halamanNecrosis I 2015Fabri CarbajalBelum ada peringkat
- Biosíntesis de La AcetilcolinaDokumen4 halamanBiosíntesis de La AcetilcolinaRomulo ChuquillanquiBelum ada peringkat
- El Metabolismo CelularDokumen14 halamanEl Metabolismo CelularHenry EnriquezBelum ada peringkat
- Logica PDFDokumen99 halamanLogica PDFLaloBelum ada peringkat
- Laboratorio 4. Citocromo OxidasaDokumen6 halamanLaboratorio 4. Citocromo OxidasaMajo ReyesBelum ada peringkat
- ADN Recombinante VHADokumen47 halamanADN Recombinante VHALuu MaidanaaBelum ada peringkat
- FARMACOCIDokumen13 halamanFARMACOCIRaquel PalominoBelum ada peringkat
- Lesion Celular y MuerteDokumen13 halamanLesion Celular y Muertemajo gsBelum ada peringkat
- VII. Aparato de GolgiDokumen12 halamanVII. Aparato de GolgiKevin JoseBelum ada peringkat
- Aparato de Golgi: Función y procesosDokumen12 halamanAparato de Golgi: Función y procesosKevin JoseBelum ada peringkat
- Aparato de GolgiDokumen12 halamanAparato de Golgiceciliaperez9751Belum ada peringkat
- 06 Bronquiolitis Aguda Viral 0Dokumen18 halaman06 Bronquiolitis Aguda Viral 0Daniel MagalaesBelum ada peringkat
- Art 8Dokumen17 halamanArt 8China LoorBelum ada peringkat
- FarmacodinamiaDokumen31 halamanFarmacodinamiaAurelia Gonzales Calle100% (1)
- Bibliografia de Pediatria y NeonatoDokumen1 halamanBibliografia de Pediatria y NeonatoAurelia Gonzales CalleBelum ada peringkat
- Microbiota normal de la uretra, vagina, conjuntiva y factores que la regulanDokumen2 halamanMicrobiota normal de la uretra, vagina, conjuntiva y factores que la regulanAurelia Gonzales CalleBelum ada peringkat
- MitosisDokumen16 halamanMitosisAurelia Gonzales CalleBelum ada peringkat
- Seminario Nro.2Dokumen49 halamanSeminario Nro.2Aurelia Gonzales CalleBelum ada peringkat
- 2 ArteriasDokumen41 halaman2 ArteriasNorita Ruiz PalaciosBelum ada peringkat
- Salud y Sociedad IIDokumen1 halamanSalud y Sociedad IIAurelia Gonzales CalleBelum ada peringkat
- MeiosisDokumen28 halamanMeiosisAurelia Gonzales CalleBelum ada peringkat
- GeneticaDokumen52 halamanGeneticaAurelia Gonzales CalleBelum ada peringkat
- TraduccionDokumen20 halamanTraduccionAurelia Gonzales CalleBelum ada peringkat
- Acidos NucleicosDokumen20 halamanAcidos NucleicosAurelia Gonzales CalleBelum ada peringkat
- ReplicacionDokumen28 halamanReplicacionAurelia Gonzales CalleBelum ada peringkat
- NucleoDokumen20 halamanNucleoAurelia Gonzales CalleBelum ada peringkat
- Tema 01 Celula Procariota y EucariotaDokumen36 halamanTema 01 Celula Procariota y EucariotaAndrea Cordova Maravi100% (1)
- Reticulo EndoplasmaticoDokumen20 halamanReticulo EndoplasmaticoAurelia Gonzales CalleBelum ada peringkat
- PeroxisomasDokumen8 halamanPeroxisomasAurelia Gonzales CalleBelum ada peringkat
- MitocondriasDokumen28 halamanMitocondriasAurelia Gonzales CalleBelum ada peringkat
- Membrana PlasmaticaDokumen22 halamanMembrana PlasmaticaAurelia Gonzales CalleBelum ada peringkat
- Tema CarbohidratosDokumen28 halamanTema CarbohidratosAurelia Gonzales CalleBelum ada peringkat
- Endosomas y LisosomasDokumen24 halamanEndosomas y LisosomasAurelia Gonzales Calle93% (14)
- TransporteDokumen32 halamanTransporteAurelia Gonzales CalleBelum ada peringkat
- Citoplasma-CitoesqueletoDokumen24 halamanCitoplasma-CitoesqueletoAurelia Gonzales Calle100% (1)
- Tema EnzimasDokumen36 halamanTema EnzimasAurelia Gonzales CalleBelum ada peringkat
- Reporte de Parasitologia Gota GruesaDokumen6 halamanReporte de Parasitologia Gota Gruesacarrascovargasjose922Belum ada peringkat
- Función pancreática LundhDokumen13 halamanFunción pancreática LundhJosue CampoverdeBelum ada peringkat
- Hemooo 12Dokumen44 halamanHemooo 12Roo DuarteBelum ada peringkat
- Informe de MiologíaDokumen32 halamanInforme de MiologíaDann LiuBelum ada peringkat
- Anatomía y fisiología de los seres vivosDokumen44 halamanAnatomía y fisiología de los seres vivosLeidy Ivonet Tintaya Yzaguirre100% (1)
- Aparato Urinario PracticaDokumen21 halamanAparato Urinario PracticaEugeo KiriyagaBelum ada peringkat
- Rúbrica Trabajo Práctico Maqueta Celula Animal 5° AñosDokumen2 halamanRúbrica Trabajo Práctico Maqueta Celula Animal 5° AñosClaudia Bastias RojasBelum ada peringkat
- Estructuras y comparación de células animales, vegetales y bacterianasDokumen6 halamanEstructuras y comparación de células animales, vegetales y bacterianasglenda bravoBelum ada peringkat
- Biología: Introducción a la ciencia de la vidaDokumen371 halamanBiología: Introducción a la ciencia de la vidaKARINA HEWITT GARCIA - EstudianteBelum ada peringkat
- Estudio histopatológico básico de la médula óseaDokumen58 halamanEstudio histopatológico básico de la médula óseaJ.V. Siritt ChangBelum ada peringkat
- ARTI I Resumen HISTOLOGIADokumen119 halamanARTI I Resumen HISTOLOGIAMicaela DuranBelum ada peringkat
- Ejercicios de Repaso Tema 1 (4º Eso)Dokumen6 halamanEjercicios de Repaso Tema 1 (4º Eso)g3x ᴅᴍᴀᴋɪɪɪBelum ada peringkat
- Sistema MuscularDokumen14 halamanSistema Muscularcamilo andres anteliz sarmientoBelum ada peringkat
- Tincion HistologiaDokumen15 halamanTincion HistologiaBárbara LeiteBelum ada peringkat
- Semana 3Dokumen17 halamanSemana 3Jaro RQBelum ada peringkat
- Tejido Sanguineo Tarea 5Dokumen4 halamanTejido Sanguineo Tarea 5Yami FretesBelum ada peringkat
- Cap. HistologíaDokumen32 halamanCap. HistologíaMauro VelascoBelum ada peringkat
- LaminasDokumen65 halamanLaminasMisc LázaroBelum ada peringkat
- Movimiento Celular Externo y LocomociónDokumen34 halamanMovimiento Celular Externo y LocomociónJorge KD0% (1)
- Tincion PapanicolauDokumen1 halamanTincion PapanicolauYuli GarcíaBelum ada peringkat
- Unab HematologiaDokumen115 halamanUnab HematologiaArianne Villagra YañezBelum ada peringkat
- Dermatología Apuntes BásicosDokumen8 halamanDermatología Apuntes BásicosAlex TuFiñoBelum ada peringkat
- Fisiologia Del EritrocitoDokumen3 halamanFisiologia Del EritrocitoOSEAS CARVAJAL CHINOBelum ada peringkat
- TINCIONESDokumen18 halamanTINCIONESMin JimenaBelum ada peringkat
- Semana 9 Músculo EsqueléticoDokumen9 halamanSemana 9 Músculo EsqueléticoAdriana Vivar Balcazar100% (1)
- Ficha Tejido OseoDokumen4 halamanFicha Tejido OseoJimmy MoralesBelum ada peringkat