Anda mungkin juga menyukai
- Organización Celular para Quinto Grado de PrimariaDokumen5 halamanOrganización Celular para Quinto Grado de PrimariaandreaBelum ada peringkat
- Protuberancia Anular o Puente de Varolio SnellDokumen6 halamanProtuberancia Anular o Puente de Varolio Snelllsantiago_293399Belum ada peringkat
- Pared de Gram NegativosDokumen31 halamanPared de Gram NegativosDarla SanchezBelum ada peringkat
- Articulaciones, Palancas y FuerzasDokumen77 halamanArticulaciones, Palancas y Fuerzasbryan trianaBelum ada peringkat
- La RespiraciónDokumen13 halamanLa RespiraciónRaquel Fernández Escobar100% (2)
- Conduccion SaltatoriaDokumen3 halamanConduccion SaltatoriaAdolfo Inda67% (3)
- Electroestimulador FinalDokumen13 halamanElectroestimulador FinalJUAN CAMILO CASALLAS FARIAS100% (1)
- Produccion Naturaleza y PropiedadesDokumen5 halamanProduccion Naturaleza y PropiedadesJazmin LafuenteBelum ada peringkat
- Formación del sistema nervioso y estructuras asociadasDokumen6 halamanFormación del sistema nervioso y estructuras asociadasGENICITA100% (1)
- Ejc 4-36 Primeros Auxilios para Caninos PDFDokumen190 halamanEjc 4-36 Primeros Auxilios para Caninos PDFS-3 BICON 50 VETAS, TIBU - NORTE DE SANTANDERBelum ada peringkat
- Glosario de AnatomiaDokumen10 halamanGlosario de AnatomiaAdi BorrayoBelum ada peringkat
- Chopin Minute Waltz Op64 No1Dokumen4 halamanChopin Minute Waltz Op64 No1Brian Leung100% (1)
- Comisuras cerebralesDokumen4 halamanComisuras cerebralesErik GarciaBelum ada peringkat
- Biomecanica de La ColumnaDokumen35 halamanBiomecanica de La ColumnaJeff VillaplanaBelum ada peringkat
- 1 Medula Espinal1Dokumen94 halaman1 Medula Espinal1Vinny ZavalaBelum ada peringkat
- Surós. Clinica Propedeutica PDFDokumen1.163 halamanSurós. Clinica Propedeutica PDFALANBelum ada peringkat
- Audición - PurvesDokumen18 halamanAudición - PurvesJochiJaramilloBelum ada peringkat
- Tejido ConectivoDokumen23 halamanTejido ConectivoDanielaRojasBelum ada peringkat
- Partes Del Sistema Respiratorio para Tercero de PrimariaDokumen4 halamanPartes Del Sistema Respiratorio para Tercero de PrimariaRossy LRBelum ada peringkat
- Articulación alveolodentaria: sus partes y funcionesDokumen19 halamanArticulación alveolodentaria: sus partes y funcionesAngie Rojas100% (1)
- Fisiopatologia Del CancerDokumen10 halamanFisiopatologia Del CancerGustavo PeñaBelum ada peringkat
- Mapa Conceptual Gonzalo Reyes Tarazona PS BARRANCA USP 8Dokumen1 halamanMapa Conceptual Gonzalo Reyes Tarazona PS BARRANCA USP 8gonzalo reyes tarazonaBelum ada peringkat
- Receptores sensoriales de la pielDokumen4 halamanReceptores sensoriales de la pielRicky Francis Gutierrez Arenas100% (1)
- Bioelectricidad: potenciales de acción y propagaciónDokumen46 halamanBioelectricidad: potenciales de acción y propagaciónFernanda AliagaBelum ada peringkat
- Resumen Tejido Conjuntivo o ConectivoDokumen7 halamanResumen Tejido Conjuntivo o ConectivoOrlandoArtemioPoloVeraBelum ada peringkat
- Vias de Conduccion NerviosaDokumen13 halamanVias de Conduccion NerviosaDANIEL ALEJANDROBelum ada peringkat
- Capitulo 8 Resumen Guyton e Hall Fisiologia Medica 13 EdDokumen8 halamanCapitulo 8 Resumen Guyton e Hall Fisiologia Medica 13 EdMinelda Meliza Cruz ChoqueBelum ada peringkat
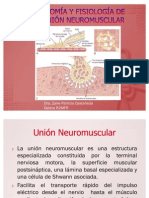
- Anatomía y Fisiología de La Unión NeuromuscularDokumen17 halamanAnatomía y Fisiología de La Unión NeuromuscularFann Ognapsy100% (1)
- CAVIDAD BUCAL (Informe)Dokumen12 halamanCAVIDAD BUCAL (Informe)Natty BD0% (1)
- Arco ReflejoDokumen10 halamanArco ReflejoBracingMeerkat Eel ChilinskyBelum ada peringkat
- Regulacion Respiración - SensoresDokumen9 halamanRegulacion Respiración - SensoresFernando Ruben Marino AguilarBelum ada peringkat
- El Sistema de HaversDokumen4 halamanEl Sistema de HaversLuis VelascoBelum ada peringkat
- Tejido Oseo y Cartilaginoso PDFDokumen63 halamanTejido Oseo y Cartilaginoso PDFAna RomeroBelum ada peringkat
- Potenciales de acción: ondas eléctricas que transmiten señales nerviosasDokumen4 halamanPotenciales de acción: ondas eléctricas que transmiten señales nerviosasJonathan MurilloBelum ada peringkat
- Musculo LisoDokumen19 halamanMusculo LisoAndrea ZepedaBelum ada peringkat
- Tejido Nervioso. HistologiaDokumen10 halamanTejido Nervioso. HistologiaManuel ChervoBelum ada peringkat
- Fisiología Del Impulso NerviosoDokumen5 halamanFisiología Del Impulso NerviosoFrancisca RiveraBelum ada peringkat
- Fracturas Del Radio y CubitoDokumen18 halamanFracturas Del Radio y CubitoIsabel María100% (1)
- Configuración interna del bulbo raquídeo y sus cortesDokumen4 halamanConfiguración interna del bulbo raquídeo y sus cortesCami GonzalezBelum ada peringkat
- Cuestionario Bulbo RaquideoDokumen2 halamanCuestionario Bulbo RaquideoLinda Zapata Elguezabal100% (1)
- La Membrana CelularDokumen23 halamanLa Membrana CelularJanet Muñoz NorambuenaBelum ada peringkat
- Propiocepcion FisiologiaDokumen20 halamanPropiocepcion FisiologiaAlexx Alvarez0% (1)
- Guyton Vias SensitivasDokumen13 halamanGuyton Vias SensitivassaraBelum ada peringkat
- Articulación Del Tobillo (Talocrural)Dokumen31 halamanArticulación Del Tobillo (Talocrural)Leandro Fiordelisi100% (1)
- Capítulo 46Dokumen26 halamanCapítulo 46Israel Arroyo Velazquez100% (4)
- Fisiología y Farmacología de La Transmisión NeuromuscularDokumen11 halamanFisiología y Farmacología de La Transmisión NeuromuscularsanjuandediosanestesiaBelum ada peringkat
- Histologia de Gandlios Sensitivo y Autonomo-1Dokumen3 halamanHistologia de Gandlios Sensitivo y Autonomo-1vivy94vcBelum ada peringkat
- Reflejo Miotatico 2020 PDFDokumen5 halamanReflejo Miotatico 2020 PDFliz0% (1)
- FX Tibia y PeronéDokumen18 halamanFX Tibia y PeronéMaximiliano Fernandez RuizBelum ada peringkat
- Neuroanatomia QuimicaDokumen15 halamanNeuroanatomia QuimicaDiego Andres Villamizar CabreraBelum ada peringkat
- Glosario de FisiologiaDokumen2 halamanGlosario de FisiologiaStephanie Gabriela CastilloBelum ada peringkat
- Clase Sobre La Histología de Los Pulmones.Dokumen2 halamanClase Sobre La Histología de Los Pulmones.elizabeth sarahy maggiorani cegarraBelum ada peringkat
- Formación del viscerocráneoDokumen11 halamanFormación del viscerocráneojoseBelum ada peringkat
- Principios Generales de La Funcion GastrointestinalDokumen34 halamanPrincipios Generales de La Funcion GastrointestinalOscar AgüeroBelum ada peringkat
- Examen EmbriologiaDokumen4 halamanExamen EmbriologiaMartha RLBelum ada peringkat
- Pasos de La Técnica Histológica BásicaDokumen2 halamanPasos de La Técnica Histológica BásicaAlejandra Ortiz FerioliBelum ada peringkat
- Transmisión neuromuscular: Unión, receptores y relajantesDokumen6 halamanTransmisión neuromuscular: Unión, receptores y relajantesHoracio Ramìrez LopezBelum ada peringkat
- Práctica Lab Medición de Líquidos in Vivo e in VitroDokumen8 halamanPráctica Lab Medición de Líquidos in Vivo e in VitroJesús MartínezBelum ada peringkat
- Tejido muscular: características y tiposDokumen5 halamanTejido muscular: características y tiposKatiAriasBelum ada peringkat
- Tejido Linfático y Órganos Linfáticos MapaDokumen10 halamanTejido Linfático y Órganos Linfáticos MapaSara OlveraBelum ada peringkat
- Potencial de AcciónDokumen16 halamanPotencial de AcciónEsteban De AbreuBelum ada peringkat
- Láminas de RexedDokumen6 halamanLáminas de RexedMiguelAngelMontañoSarmientoBelum ada peringkat
- Mecanoreceptores PDFDokumen9 halamanMecanoreceptores PDFrfenoglioBelum ada peringkat
- Factor de Seguridad para La Transmisión en La Unión NeuromuscularDokumen1 halamanFactor de Seguridad para La Transmisión en La Unión NeuromuscularlorenBelum ada peringkat
- Contraccion MuscularDokumen8 halamanContraccion MuscularAngel LópezBelum ada peringkat
- Via OlfatoriaDokumen11 halamanVia OlfatoriaANA ESPINOSABelum ada peringkat
- 3- Potenciales de Membrana y Potenciales de AcciónDokumen63 halaman3- Potenciales de Membrana y Potenciales de Accióndr.josefernandezlBelum ada peringkat
- Plan de Clase Practicas IV - Enero 2019 - 2Dokumen9 halamanPlan de Clase Practicas IV - Enero 2019 - 2Gustavo PeñaBelum ada peringkat
- Determinación de Extracto Etéreo en Leche enDokumen3 halamanDeterminación de Extracto Etéreo en Leche enDennysFabriBelum ada peringkat
- DDokumen38 halamanDLuis Felipe RamirezBelum ada peringkat
- 2 2 Tecnicas Adm VacDokumen6 halaman2 2 Tecnicas Adm VacKarolina IntriagoBelum ada peringkat
- DROGAS NEFROTÓXICAS-ExposicDokumen6 halamanDROGAS NEFROTÓXICAS-ExposicGian Carlos MedinaBelum ada peringkat
- Intervalos de Confianza EjemploDokumen7 halamanIntervalos de Confianza EjemploGustavo PeñaBelum ada peringkat
- Casella - Five Easy Pieces 'Pupazzetti' 4Dokumen36 halamanCasella - Five Easy Pieces 'Pupazzetti' 4Gustavo PeñaBelum ada peringkat
- Huasca de OcampoDokumen23 halamanHuasca de OcampoGustavo PeñaBelum ada peringkat
- Nefrotico Caso PDFDokumen4 halamanNefrotico Caso PDFGustavo PeñaBelum ada peringkat
- VigorexiaDokumen18 halamanVigorexiaGustavo PeñaBelum ada peringkat
- INTERPRETACION - Indicadores Cresimiento InfantelDokumen56 halamanINTERPRETACION - Indicadores Cresimiento InfantelJuan MirandaBelum ada peringkat
- Cronología de La ErupciónDokumen2 halamanCronología de La ErupciónGustavo PeñaBelum ada peringkat
- El Alquimista ResumenDokumen5 halamanEl Alquimista ResumenGustavo PeñaBelum ada peringkat
- Protocolo de Atencion A La Mujer GetanteDokumen42 halamanProtocolo de Atencion A La Mujer GetanteGustavo PeñaBelum ada peringkat
- PLAQUETASDokumen12 halamanPLAQUETASGustavo PeñaBelum ada peringkat
- Medidas de Tendencia CentralDokumen2 halamanMedidas de Tendencia CentralGustavo PeñaBelum ada peringkat
- JacobDokumen1 halamanJacobGustavo PeñaBelum ada peringkat
- Mujer Embarazada en El Consul To RioDokumen5 halamanMujer Embarazada en El Consul To RiopactyBelum ada peringkat
- Uniones Celulares 2008Dokumen12 halamanUniones Celulares 2008riesgos33Belum ada peringkat
- Mico BacteriasDokumen20 halamanMico BacteriasGustavo PeñaBelum ada peringkat
- MosiahDokumen1 halamanMosiahGustavo PeñaBelum ada peringkat
- Triptico Ciencias NaturalesDokumen1 halamanTriptico Ciencias NaturalesGustavo PeñaBelum ada peringkat
- Aparato Masticatorio CSODokumen15 halamanAparato Masticatorio CSOsilviasc_5475% (4)
- Nom 010 Ssa2 2010Dokumen13 halamanNom 010 Ssa2 2010Gustavo PeñaBelum ada peringkat
- Factores de RiesgoDokumen1 halamanFactores de RiesgoGustavo PeñaBelum ada peringkat
- Estructuras de Los 20 Aminoácidos ComunesDokumen9 halamanEstructuras de Los 20 Aminoácidos ComunesGustavo PeñaBelum ada peringkat
- Fisiologia Tema 3 OseoDokumen16 halamanFisiologia Tema 3 OseobernyBelum ada peringkat
- Dolor de mamas: causas, tipos y tratamientoDokumen4 halamanDolor de mamas: causas, tipos y tratamientoAni OrtegaBelum ada peringkat
- Trabajo de LigamentosDokumen46 halamanTrabajo de LigamentosEdgardo Manuel Uchuya ChavezBelum ada peringkat
- Piel ResumenDokumen4 halamanPiel ResumenMaría PazBelum ada peringkat
- Insuficiencia Renal Aguda Fin 2Dokumen21 halamanInsuficiencia Renal Aguda Fin 2Merly Lizeth Contreras InfantesBelum ada peringkat
- El Sistema OseoDokumen1 halamanEl Sistema OseoMiguel ChoBelum ada peringkat
- Embriología Del Aparato DigestivoDokumen2 halamanEmbriología Del Aparato DigestivoKarina Hernàndez Blanco100% (1)
- Glándulas endocrinas y criminalidadDokumen3 halamanGlándulas endocrinas y criminalidadConcepcion PeraltaBelum ada peringkat
- Anatomía y Fisiología de La BocaDokumen6 halamanAnatomía y Fisiología de La BocaAdriianiita HoffmanBelum ada peringkat
- Anatomía de OidoDokumen37 halamanAnatomía de OidoBenjamin Misael Rivera DiazBelum ada peringkat
- Orientacion de Esofago, Estomao, Duodeno YeyunoDokumen18 halamanOrientacion de Esofago, Estomao, Duodeno YeyunoSarah duranBelum ada peringkat
- Ancylostoma CaninumDokumen3 halamanAncylostoma Caninumigdamaco0% (1)
- Sistema Digestivo de Los MonogastricosDokumen2 halamanSistema Digestivo de Los MonogastricosestefanimrojasBelum ada peringkat
- Cap 054Dokumen4 halamanCap 054Yuri medranoBelum ada peringkat
- Desarrollo InmunitarioDokumen6 halamanDesarrollo InmunitarioBrian Jhesit GarciaBelum ada peringkat
- 04.aparato Reproductor Femenino PDFDokumen99 halaman04.aparato Reproductor Femenino PDFRoberto SantacruzBelum ada peringkat
- La médula ósea, la sangre y las células sanguíneasDokumen33 halamanLa médula ósea, la sangre y las células sanguíneasjulio cesarBelum ada peringkat
- El Sistema de Meridianos y ColateralesDokumen6 halamanEl Sistema de Meridianos y ColateralesHerman H. FloresBelum ada peringkat
- Circulación cerebral: transporte vital para el cerebroDokumen15 halamanCirculación cerebral: transporte vital para el cerebroJunior David Cruz CeliBelum ada peringkat
- Clase 8. Analisis de Enfermedades Neurovegetativas Ligadas A Estructuras EstudiadasDokumen31 halamanClase 8. Analisis de Enfermedades Neurovegetativas Ligadas A Estructuras EstudiadasmmercedestcBelum ada peringkat
- Pares CranealesDokumen5 halamanPares CranealesGabriela Mendoza MendozaBelum ada peringkat
- Guia FisioDokumen24 halamanGuia FisioEdher ArturoBelum ada peringkat
- Trabajo Practico Sistema DigestivoDokumen2 halamanTrabajo Practico Sistema DigestivoMonica Brezovec0% (1)
- Laboratorio de hormonas: aspectos prácticos en endocrinologíaDokumen13 halamanLaboratorio de hormonas: aspectos prácticos en endocrinologíaAngel Agustin Marinez HernandezBelum ada peringkat
- 1120202862.apunte Anatomia y Fisiologia - 2018Dokumen27 halaman1120202862.apunte Anatomia y Fisiologia - 2018josueBelum ada peringkat