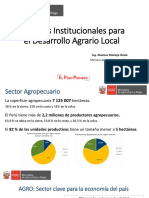
Anda mungkin juga menyukai
- ForespiracionDokumen19 halamanForespiracionAaron MelendresBelum ada peringkat
- Clase 5 AcodosDokumen4 halamanClase 5 AcodosANGEL PEDRO CAMPOS JULCABelum ada peringkat
- Informe Tecnica de Comunicacion Maiz Duro y DulceDokumen4 halamanInforme Tecnica de Comunicacion Maiz Duro y DulceJose Luis Rodriguez NiveloBelum ada peringkat
- Fisiología de CultivosDokumen13 halamanFisiología de CultivosDiana Carolina Legarda LopezBelum ada peringkat
- Mejoramiento Genético Del ArrozDokumen17 halamanMejoramiento Genético Del ArrozJosé Angel VázquezBelum ada peringkat
- Modulo6 ArvensesDokumen30 halamanModulo6 Arvensesdiego vargasBelum ada peringkat
- Árboles de Alto RendimientoDokumen32 halamanÁrboles de Alto Rendimientoamparo vidales100% (1)
- Plantas c4Dokumen18 halamanPlantas c4anon_797932374Belum ada peringkat
- TriacontanolDokumen32 halamanTriacontanolcristiam leguia yaurisBelum ada peringkat
- Variedades y Mejoramiento Genetico Del CafeDokumen5 halamanVariedades y Mejoramiento Genetico Del CafeGamus del PeruBelum ada peringkat
- Área FoliarDokumen10 halamanÁrea Foliarbelen sofiBelum ada peringkat
- Analisis de Crecimiento Del Pasto Maralfalfa en Clima Cálido Subhúmedo.Dokumen14 halamanAnalisis de Crecimiento Del Pasto Maralfalfa en Clima Cálido Subhúmedo.Raul KimBelum ada peringkat
- Fisioloía Reporte No.5Dokumen8 halamanFisioloía Reporte No.5Erick DelgadoBelum ada peringkat
- Informe de Invernadero de RosasDokumen14 halamanInforme de Invernadero de RosasFranz omar Jamanca GonzalesBelum ada peringkat
- Vasquez Libro 2017Dokumen83 halamanVasquez Libro 2017Alonzo EspinozaBelum ada peringkat
- Biotecnologia TomateDokumen26 halamanBiotecnologia TomateRICHARD R.Belum ada peringkat
- Aplicacion y EvaluacionDokumen32 halamanAplicacion y EvaluacionJosé Mario Valle PuenteBelum ada peringkat
- Fisiología de Plantas en Condiciones de EstrésDokumen21 halamanFisiología de Plantas en Condiciones de EstrésJeanpieer BG100% (1)
- Piña InformeDokumen11 halamanPiña InformegloriaBelum ada peringkat
- FOTORESPIRACIONDokumen12 halamanFOTORESPIRACIONerafaelpaBelum ada peringkat
- Ensayo FotorespiraciónDokumen8 halamanEnsayo FotorespiraciónCarito Lisseth BorjaBelum ada peringkat
- Maiz - Fertilizacion PDFDokumen5 halamanMaiz - Fertilizacion PDFroosvelt vasquez chicomaBelum ada peringkat
- Fitomejoramiento 1 UNADDokumen16 halamanFitomejoramiento 1 UNADAndrésKoflaBelum ada peringkat
- HD Dormición de SemillasDokumen20 halamanHD Dormición de SemillasPrimero De AgropecuariasBelum ada peringkat
- Etapa BioquimicaDokumen42 halamanEtapa BioquimicaAntonio OmogrossoBelum ada peringkat
- Cultivo de TejidoDokumen35 halamanCultivo de TejidoEnrique Ruiz TapiaBelum ada peringkat
- AngiospermaeDokumen14 halamanAngiospermaeDavid DiazBelum ada peringkat
- Informe 5Dokumen10 halamanInforme 5Marcos Beckham Lino MontesBelum ada peringkat
- Reporte Final Ecologia de MalezasDokumen17 halamanReporte Final Ecologia de MalezasDennis Najera100% (1)
- Proyecto BiotecnologiaDokumen30 halamanProyecto BiotecnologiaazucenaBelum ada peringkat
- Otra 612 CalabacínDokumen20 halamanOtra 612 CalabacínSergi PillacaBelum ada peringkat
- Plantas c3 y c4 FinalDokumen11 halamanPlantas c3 y c4 FinalROSA MARIA GIANELLA REATEGUI SIANCASBelum ada peringkat
- 8 Abonos Verdes PDFDokumen23 halaman8 Abonos Verdes PDFJorge CarrilloBelum ada peringkat
- CebollaDokumen11 halamanCebollaJosé Corona100% (1)
- NEMATODODokumen19 halamanNEMATODOedilBelum ada peringkat
- El Combate de MalezasDokumen19 halamanEl Combate de MalezasNeiver Jesús Flores MenorBelum ada peringkat
- Fertilización FoliarDokumen21 halamanFertilización FoliarAldo Leopoldo MottoBelum ada peringkat
- Clasificacion Hortalizas Ago 16Dokumen11 halamanClasificacion Hortalizas Ago 16Jesus MartinezBelum ada peringkat
- Como Criar Mariposas PDFDokumen2 halamanComo Criar Mariposas PDFKristenBelum ada peringkat
- Virus Del Mosaico de La Alfalfa YaDokumen16 halamanVirus Del Mosaico de La Alfalfa Yajulian rojas s.Belum ada peringkat
- Ciat HibridacionDokumen41 halamanCiat HibridacionSERGIOBelum ada peringkat
- Potencial Hidrico de Solanum TuberosumDokumen11 halamanPotencial Hidrico de Solanum TuberosumAlexis Díaz56% (9)
- Tratamiento PregerminativoDokumen18 halamanTratamiento PregerminativosantiagoBelum ada peringkat
- Absorcion de Nutrientes PDFDokumen29 halamanAbsorcion de Nutrientes PDFWilliam Yucra RodasBelum ada peringkat
- Plagas Insecto en PapaDokumen27 halamanPlagas Insecto en PapaJulissa Espino GonzalesBelum ada peringkat
- Examen-Fundamentos de Fitotecnia PDFDokumen5 halamanExamen-Fundamentos de Fitotecnia PDFyakkito_chantaBelum ada peringkat
- Empleo de Hormonas (ANA y AIB) Estimuladoras Del Enraizamiento para La Propagación Vegetativa de CHLDokumen5 halamanEmpleo de Hormonas (ANA y AIB) Estimuladoras Del Enraizamiento para La Propagación Vegetativa de CHLNyxbillcito MeandyouBelum ada peringkat
- MUSLE PracticaDokumen88 halamanMUSLE PracticaCintia RamosBelum ada peringkat
- CULTIVO DE AJI PAPRIKADokumen31 halamanCULTIVO DE AJI PAPRIKADavid Javier Garcia Silvestre100% (1)
- Guía Del Herbario. Universidad Rafael LandivarDokumen17 halamanGuía Del Herbario. Universidad Rafael LandivarHarold PérezBelum ada peringkat
- Obtención de plantas de palma datilera mediante germinación de semillas in vitro con citoquininasDokumen7 halamanObtención de plantas de palma datilera mediante germinación de semillas in vitro con citoquininasAmerica CorreaBelum ada peringkat
- Encalado CafeDokumen7 halamanEncalado CafeDavid GomezBelum ada peringkat
- Heladas agrícolasDokumen33 halamanHeladas agrícolasAlejandra RamírezBelum ada peringkat
- ChardakovDokumen5 halamanChardakovAndreaForteRuizBelum ada peringkat
- Curso Agroclimatologia Aplicada A Pequenos ProductoresDokumen147 halamanCurso Agroclimatologia Aplicada A Pequenos Productoresalejandro100% (1)
- Tarea 3 - Clasificacion - Botanica - Taxonomica Diego Fernando TorraDokumen8 halamanTarea 3 - Clasificacion - Botanica - Taxonomica Diego Fernando Torradiego fernando torra cañasBelum ada peringkat
- Manejo Del Picudo Negro Del PlatanoDokumen15 halamanManejo Del Picudo Negro Del PlatanoDaniel Marín100% (1)
- Examen Final Fruti GeneralDokumen5 halamanExamen Final Fruti GeneralElmer Isaac Tongo BrionesBelum ada peringkat
- Manejo integrado de la polilla guatemalteca de la papa (Tecia solanivora, Povolny)Dari EverandManejo integrado de la polilla guatemalteca de la papa (Tecia solanivora, Povolny)Belum ada peringkat
- Manejo fitosanitario del cultivo de gulupa (Passiflora edulis Sims.): Medidas para la temporada invernalDari EverandManejo fitosanitario del cultivo de gulupa (Passiflora edulis Sims.): Medidas para la temporada invernalBelum ada peringkat
- Manejo Integrado en La Produccion y Sanidad Del ArrozDokumen40 halamanManejo Integrado en La Produccion y Sanidad Del ArrozRenzo FernandezBelum ada peringkat
- Capitulo IIDokumen25 halamanCapitulo IIAriana PuiconBelum ada peringkat
- 2020 Propuesta Mejora Proceso Teses EmpaquesDokumen55 halaman2020 Propuesta Mejora Proceso Teses EmpaquesEstefania Cajas OñaBelum ada peringkat
- 2 - MinagriDokumen19 halaman2 - MinagriAriana PuiconBelum ada peringkat
- Clase N°3 Calidad y BpaDokumen26 halamanClase N°3 Calidad y BpaAriana PuiconBelum ada peringkat
- Degradacion Del SueloDokumen1 halamanDegradacion Del SueloAriana PuiconBelum ada peringkat
- Cadena Productiva de Arveja y Haba: Una Experiencia en Acobamba-HuancavelicaDokumen100 halamanCadena Productiva de Arveja y Haba: Una Experiencia en Acobamba-HuancavelicacareperuBelum ada peringkat
- Banco de Preguntas de PoscosechaDokumen9 halamanBanco de Preguntas de PoscosechaAriana PuiconBelum ada peringkat
- TC104104 PDFDokumen80 halamanTC104104 PDFAriana PuiconBelum ada peringkat
- Manual Agrotecnia 02Dokumen286 halamanManual Agrotecnia 02Ariana PuiconBelum ada peringkat
- Cadena de FrioDokumen51 halamanCadena de FrioJosé Castro0% (1)
- Biotecnologia en La Salud PDFDokumen56 halamanBiotecnologia en La Salud PDFprincipelicBelum ada peringkat
- Haccp PDFDokumen162 halamanHaccp PDFAriana PuiconBelum ada peringkat
- Buenasprcticasdemanufactura 110816190224 Phpapp02Dokumen33 halamanBuenasprcticasdemanufactura 110816190224 Phpapp02kary2806Belum ada peringkat
- Tesis en Cebolla PDFDokumen72 halamanTesis en Cebolla PDFAriana PuiconBelum ada peringkat
- Condiciones laborales de los trabajadores de maestranza en un hospital públicoDokumen114 halamanCondiciones laborales de los trabajadores de maestranza en un hospital públicoAriana PuiconBelum ada peringkat
- Cultivo de Anteras Ciat PDFDokumen1 halamanCultivo de Anteras Ciat PDFAriana PuiconBelum ada peringkat
- Sanchez Carmen Siniestralidad Laboral Sector ConstruccionDokumen165 halamanSanchez Carmen Siniestralidad Laboral Sector ConstruccionErik PattersonBelum ada peringkat
- Enfermedades Del Cultivo de PiñaDokumen11 halamanEnfermedades Del Cultivo de PiñaAriana PuiconBelum ada peringkat
- Proyecto Integrador Saberes 161020234545 PDFDokumen66 halamanProyecto Integrador Saberes 161020234545 PDFAriana PuiconBelum ada peringkat
- Temperatura PDFDokumen5 halamanTemperatura PDFFernando GarciaBelum ada peringkat
- Temperatura PDFDokumen5 halamanTemperatura PDFFernando GarciaBelum ada peringkat
- Encuesta Clima Laboral GC PDFDokumen6 halamanEncuesta Clima Laboral GC PDFAriana PuiconBelum ada peringkat
- Encuesta Clima Laboral GC PDFDokumen6 halamanEncuesta Clima Laboral GC PDFAriana PuiconBelum ada peringkat
- Formulario Clima OrganizacionalDokumen6 halamanFormulario Clima OrganizacionalenjoydabookBelum ada peringkat
- Fitopatología Del Cultivo de PapaDokumen31 halamanFitopatología Del Cultivo de PapaAriana PuiconBelum ada peringkat
- Encuesta Clima Laboral GC PDFDokumen6 halamanEncuesta Clima Laboral GC PDFAriana PuiconBelum ada peringkat
- Condiciones laborales de los trabajadores de maestranza en un hospital públicoDokumen114 halamanCondiciones laborales de los trabajadores de maestranza en un hospital públicoAriana PuiconBelum ada peringkat
- Manual Condiciones y Medio Ambient eDokumen78 halamanManual Condiciones y Medio Ambient eLuis Gabriel Alcócer MonteroBelum ada peringkat
- Grupo 4Dokumen22 halamanGrupo 4Ylse LorenaBelum ada peringkat
- Nube Minera Planificacion Open Pit Seleccion Del Metodo de ExplotacionDokumen61 halamanNube Minera Planificacion Open Pit Seleccion Del Metodo de ExplotacionRodrigo Sánchez FazioBelum ada peringkat
- Hidrografía de GuatemalaDokumen30 halamanHidrografía de GuatemalaMisael Ebt0% (2)
- Plantilla RestauranteDokumen1 halamanPlantilla RestauranteSILVIA PATRICIA OSORIO CANTILLOBelum ada peringkat
- Regreso A La Nueva Normalidad Prepa Ibero PDFDokumen1 halamanRegreso A La Nueva Normalidad Prepa Ibero PDFBetty Ramírez EscobedoBelum ada peringkat
- Universidad Francisco de Paula Santander Ocaña: Autores Facultad Plan de Estudios Director Título de La TesisDokumen223 halamanUniversidad Francisco de Paula Santander Ocaña: Autores Facultad Plan de Estudios Director Título de La TesishitaBelum ada peringkat
- Requerimientos de ObraDokumen18 halamanRequerimientos de ObraAC ArceliBelum ada peringkat
- Equilibrio QuimicoDokumen8 halamanEquilibrio QuimicoDennis LinoBelum ada peringkat
- Actividad 1 Gestion AmbientalDokumen1 halamanActividad 1 Gestion AmbientalJuan Victor Santa CruzBelum ada peringkat
- Taller MedicamentosDokumen2 halamanTaller MedicamentospacoBelum ada peringkat
- Clase 4. PerioperatorioDokumen32 halamanClase 4. Perioperatorioblanca ortegaBelum ada peringkat
- Fulguraciones Solares y Sus Efectos 1Dokumen12 halamanFulguraciones Solares y Sus Efectos 1Luis Felipe Cazares HernandezBelum ada peringkat
- El Sistema CirculatorioDokumen11 halamanEl Sistema CirculatorioMarce AlbarracinBelum ada peringkat
- Generalidades de La Ecografia ObstetricaDokumen60 halamanGeneralidades de La Ecografia ObstetricaJhosy Portuguez GomezBelum ada peringkat
- Criminología Del DesarrolloDokumen15 halamanCriminología Del DesarrolloMtra Karla LeonBelum ada peringkat
- Triptico QuemadurasDokumen2 halamanTriptico QuemadurasNORMA GONZALEZBelum ada peringkat
- Delmastic 086 Hs - Mastic 86% SólidosDokumen3 halamanDelmastic 086 Hs - Mastic 86% SólidosJavier Michel Esteban ViñegasBelum ada peringkat
- Instrucciones Autoinyeccion Sayana PressDokumen5 halamanInstrucciones Autoinyeccion Sayana PressNeyda GuevaraBelum ada peringkat
- Atlas Copco Boomer InstruccionesDokumen90 halamanAtlas Copco Boomer InstruccionesMiguelBelum ada peringkat
- RosáceaDokumen5 halamanRosáceaSofia Vela PBelum ada peringkat
- W Final Del TepsiDokumen9 halamanW Final Del TepsiBerenisce Gonzales GameroBelum ada peringkat
- Investigación del Tardígrado: Un Animal ResistenteDokumen6 halamanInvestigación del Tardígrado: Un Animal ResistenteLuis Antony Sandoval DamianBelum ada peringkat
- Formato para El Análisis de La Problemática. Tarea 3 ÉticaDokumen6 halamanFormato para El Análisis de La Problemática. Tarea 3 ÉticaTatiana VillamizarBelum ada peringkat
- Productos Rabago 2016 R1, R2 y R8 TalentuaDokumen39 halamanProductos Rabago 2016 R1, R2 y R8 TalentuaCristie MtzBelum ada peringkat
- Agroecosistema 3Dokumen11 halamanAgroecosistema 3maday camposBelum ada peringkat
- Análisis BufferDokumen26 halamanAnálisis Bufferkarin santosBelum ada peringkat
- CIED PDVSA - Fluidos de PerforaciónDokumen285 halamanCIED PDVSA - Fluidos de PerforaciónCarlos Chacon60% (5)
- Grout MorteroDokumen2 halamanGrout MorteroRichard GarciaBelum ada peringkat
- Medina-1977 Introduccion A La Ecofisiologia Vegetal PDFDokumen112 halamanMedina-1977 Introduccion A La Ecofisiologia Vegetal PDFCarlos Andres RodriguezBelum ada peringkat
- Taller RCVDokumen7 halamanTaller RCVNicol AbreoBelum ada peringkat