Anda mungkin juga menyukai
- Module 3 Metabolism of Carbohydrates, LipidsDokumen209 halamanModule 3 Metabolism of Carbohydrates, Lipidsninas1112Belum ada peringkat
- Selected Topics in the History of Biochemistry. Personal Recollections. Part IIIDari EverandSelected Topics in the History of Biochemistry. Personal Recollections. Part IIIPenilaian: 1 dari 5 bintang1/5 (1)
- Clinical Biochemistry V3: Contemporary Theories and TechniquesDari EverandClinical Biochemistry V3: Contemporary Theories and TechniquesHerbert SpiegelBelum ada peringkat
- Metabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaDokumen60 halamanMetabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaAmanda PutriBelum ada peringkat
- B Y: - Idr. Megha Gaur BDS IDokumen89 halamanB Y: - Idr. Megha Gaur BDS IRishab GaurBelum ada peringkat
- Sphingolipids: Dr. Abir Alghanouchi Biochemistry Department Sciences CollegeDokumen12 halamanSphingolipids: Dr. Abir Alghanouchi Biochemistry Department Sciences CollegeUzac BenuBelum ada peringkat
- General Introduction and Basic Biochemical PrinciplesDokumen6 halamanGeneral Introduction and Basic Biochemical PrinciplescelecosibBelum ada peringkat
- Fatty Acid Synthesis 11.12.19Dokumen18 halamanFatty Acid Synthesis 11.12.19Sanreet Randhawa100% (1)
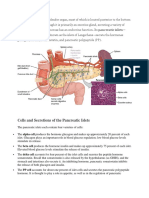
- Cells and Secretions of The Pancreatic IsletsDokumen4 halamanCells and Secretions of The Pancreatic IsletsSophia OcayBelum ada peringkat
- Carbohydrates SummaryDokumen9 halamanCarbohydrates SummaryHarold NagunaBelum ada peringkat
- l08 Metabolism in Liver 2 - Fatty Acid Metabolism (Mod)Dokumen21 halamanl08 Metabolism in Liver 2 - Fatty Acid Metabolism (Mod)Swapnil100% (4)
- Chapter 2 P-2 Enzyme-Inhibition 1Dokumen39 halamanChapter 2 P-2 Enzyme-Inhibition 1Raihan I. SakibBelum ada peringkat
- Biomolecules: Amino Acids, Peptides, and ProteinsDokumen55 halamanBiomolecules: Amino Acids, Peptides, and ProteinsBoooooooseBelum ada peringkat
- S13.CHO .MetabolismDokumen18 halamanS13.CHO .MetabolismGhea Jovita SinagaBelum ada peringkat
- PROTEIN METABOLISM Dea Farha Fira Darson FineDokumen44 halamanPROTEIN METABOLISM Dea Farha Fira Darson FineFarhati MardhiyahBelum ada peringkat
- LipidsDokumen13 halamanLipidsalianaBelum ada peringkat
- Mode of Action: InsulinDokumen8 halamanMode of Action: Insulinmanus7777100% (1)
- 2 - Lipid Metabolism Lecture For StudentsDokumen68 halaman2 - Lipid Metabolism Lecture For StudentshwhsgxBelum ada peringkat
- Digestion and Absorption of Carbohydrates: by Arpitha 3 BnysDokumen31 halamanDigestion and Absorption of Carbohydrates: by Arpitha 3 BnysBig ZeroBelum ada peringkat
- Nucleic Acid MetabolismDokumen35 halamanNucleic Acid MetabolismAmreen KhanBelum ada peringkat
- Clinical SignificanceDokumen31 halamanClinical Significancejav israelBelum ada peringkat
- Biochemistry (GREEN)Dokumen18 halamanBiochemistry (GREEN)BRYAN BALDOMEROBelum ada peringkat
- OrabyBiochemistry P.2 PDFDokumen232 halamanOrabyBiochemistry P.2 PDFJayadev Ch100% (1)
- Metabolism of CarbohydrateDokumen48 halamanMetabolism of CarbohydrateAbdullah TheBelum ada peringkat
- ACFrOgB4ugyviBu1XK4Lh1UX8Pt64wVQwa2 Exi6l8nAFhE Uu1QLN5OIKowfbqMbZ48dVUAL2yq7eDi4HBBlOHGoCWigzrxxjl305MoZfTvJxd54XHQjc6yi-YzD8c PDFDokumen12 halamanACFrOgB4ugyviBu1XK4Lh1UX8Pt64wVQwa2 Exi6l8nAFhE Uu1QLN5OIKowfbqMbZ48dVUAL2yq7eDi4HBBlOHGoCWigzrxxjl305MoZfTvJxd54XHQjc6yi-YzD8c PDFLpBelum ada peringkat
- Respiratory Chain & Oxidative PhosphorylationDokumen57 halamanRespiratory Chain & Oxidative PhosphorylationHanifa AffianiBelum ada peringkat
- PDH Complex REVIEW: DR Navin SDokumen23 halamanPDH Complex REVIEW: DR Navin Svaseem1981Belum ada peringkat
- Practical BiochemistryDokumen35 halamanPractical BiochemistryMockinjay100% (1)
- Hormones and Metabolic RegulationDokumen40 halamanHormones and Metabolic RegulationEka HabinaBelum ada peringkat
- Biochemistry of Digestive SystemDokumen55 halamanBiochemistry of Digestive SystemSyam UnhasBelum ada peringkat
- Git Biochemistry of GitDokumen9 halamanGit Biochemistry of GitSibatBelum ada peringkat
- Biochem HomeworkDokumen13 halamanBiochem Homeworkfcukingfranztastik50% (2)
- Case StudyDokumen1 halamanCase Studyapi-311718939Belum ada peringkat
- Protein TargettingDokumen11 halamanProtein TargettingPMIB Matrikulasi FKUI 2018/2019Belum ada peringkat
- Biochemistry One LinersDokumen11 halamanBiochemistry One Linerssandeep MahapatraBelum ada peringkat
- Physiology. Hrd.Dokumen549 halamanPhysiology. Hrd.Sʌɩĸʌt PʌʋɭBelum ada peringkat
- Concise Biochemistry: Fundamental Principles: March 2016Dokumen52 halamanConcise Biochemistry: Fundamental Principles: March 2016Sagar DeshmaniBelum ada peringkat
- Fatty Acid OxidationDokumen28 halamanFatty Acid OxidationEmm NomanBelum ada peringkat
- Biochemistry Lecture 2 Cell and OrganellesDokumen16 halamanBiochemistry Lecture 2 Cell and OrganellesProfessor Rakesh Sharma Biochemistry LecturesBelum ada peringkat
- Lecture 6 (Bioenergetics of Physical Exercise)Dokumen39 halamanLecture 6 (Bioenergetics of Physical Exercise)Helmi RaisBelum ada peringkat
- CH 19 Integration of MetabolismDokumen63 halamanCH 19 Integration of Metabolismshee140802100% (1)
- 3 LipidsDokumen44 halaman3 LipidsSuresh ChovatiyaBelum ada peringkat
- Metabolism of LipidsDokumen47 halamanMetabolism of LipidsMarwahBelum ada peringkat
- Hexose Monophosphate ShuntDokumen43 halamanHexose Monophosphate ShuntSecret AgentBelum ada peringkat
- Porphyrias and Errors in Heme Metabolism: Rakesh Sharma, PH.DDokumen33 halamanPorphyrias and Errors in Heme Metabolism: Rakesh Sharma, PH.DProfessor Rakesh Sharma Biochemistry Lectures0% (1)
- Complex Carbohydrates - GlycoproteinsDokumen28 halamanComplex Carbohydrates - GlycoproteinsJessica SannohBelum ada peringkat
- Citric Acid Cycle - Pyruvate DehydrogenaseDokumen33 halamanCitric Acid Cycle - Pyruvate Dehydrogenasesultan khabeeb100% (1)
- MCQ MembranesDokumen10 halamanMCQ MembranesMarilyne RizkBelum ada peringkat
- Lipoprotein MetabolismDokumen23 halamanLipoprotein MetabolismDarien LiewBelum ada peringkat
- 4 Bioenergetics and Oxidative Metabolism IIDokumen3 halaman4 Bioenergetics and Oxidative Metabolism IILinus LiuBelum ada peringkat
- Glucose, Part1Dokumen33 halamanGlucose, Part1SarahBelum ada peringkat
- Biochemistry VivaDokumen8 halamanBiochemistry VivaNazia WasimBelum ada peringkat
- Integration of Metabolism Integration of MetabolismDokumen10 halamanIntegration of Metabolism Integration of MetabolismEdison LucianoBelum ada peringkat
- Chapter 2 - Metabolism & Bioenergetics (Part 2) PDFDokumen69 halamanChapter 2 - Metabolism & Bioenergetics (Part 2) PDFdarren100% (2)
- Starch and Sucrose MetabolismDokumen41 halamanStarch and Sucrose MetabolismSanaur Rahman0% (1)
- CARBOHYDRATES With Case StudyDokumen1 halamanCARBOHYDRATES With Case StudyGeraldine Roxas CruzBelum ada peringkat
- Biochemistry Notes For BoardsDokumen35 halamanBiochemistry Notes For BoardsUjjwal PyakurelBelum ada peringkat
- Fatty Acid MetabolismDokumen40 halamanFatty Acid MetabolismMichelle Arredondo100% (1)
- Phenylalanine and Tyrosine Metabolism (18 Oct)Dokumen14 halamanPhenylalanine and Tyrosine Metabolism (18 Oct)api-19824406100% (6)
- 15 - Metabolism of Other HexosesDokumen14 halaman15 - Metabolism of Other HexosescheckmateBelum ada peringkat
- PedoDokumen2 halamanPedocheckmateBelum ada peringkat
- 18 - Lipid MetabolismDokumen26 halaman18 - Lipid Metabolismcheckmate100% (1)
- 19 - Lipid MetabolismDokumen35 halaman19 - Lipid MetabolismcheckmateBelum ada peringkat
- Lipid Metabolism - I: Chemistry, Digestion and Absorption of LipidsDokumen38 halamanLipid Metabolism - I: Chemistry, Digestion and Absorption of LipidscheckmateBelum ada peringkat
- OrthoDokumen2 halamanOrthocheckmateBelum ada peringkat
- 9 - Water Soluble VitaminsDokumen43 halaman9 - Water Soluble VitaminscheckmateBelum ada peringkat
- 13 - GluconeogenesisDokumen23 halaman13 - GluconeogenesischeckmateBelum ada peringkat
- 15 - Metabolism of Other HexosesDokumen14 halaman15 - Metabolism of Other HexosescheckmateBelum ada peringkat
- 11 - Carbohydrate MetabolismDokumen68 halaman11 - Carbohydrate MetabolismcheckmateBelum ada peringkat
- 12 - Glycogen MetabolismDokumen34 halaman12 - Glycogen Metabolismcheckmate100% (1)
- Cranial Nerves: MR Christopher Simon BSC (Neuroscience / Cognitive Psychology) MSC (Human Anatomy)Dokumen66 halamanCranial Nerves: MR Christopher Simon BSC (Neuroscience / Cognitive Psychology) MSC (Human Anatomy)checkmateBelum ada peringkat
- 14 - Blood Glucose HomeostasisDokumen34 halaman14 - Blood Glucose HomeostasischeckmateBelum ada peringkat
- 10 - Water Soluble VitaminsDokumen48 halaman10 - Water Soluble VitaminscheckmateBelum ada peringkat
- Lecture - 2 Transport Processes: Dr. S. Annie Jeyachristy Lecturer, Unit of Biochemistry Faculty of MedicineDokumen9 halamanLecture - 2 Transport Processes: Dr. S. Annie Jeyachristy Lecturer, Unit of Biochemistry Faculty of MedicinecheckmateBelum ada peringkat
- 7 - EnzymesDokumen15 halaman7 - EnzymescheckmateBelum ada peringkat
- 8 - Fat Soluble VitaminsDokumen49 halaman8 - Fat Soluble VitaminscheckmateBelum ada peringkat
- 4 - EnzymesDokumen31 halaman4 - EnzymescheckmateBelum ada peringkat
- 5 - EnzymesDokumen23 halaman5 - EnzymescheckmateBelum ada peringkat
- 6 - EnzymesDokumen13 halaman6 - EnzymescheckmateBelum ada peringkat
- Introduction To AnatomyDokumen54 halamanIntroduction To AnatomycheckmateBelum ada peringkat
- Embryology of Cardiovascular System 1: DR - Navajyothi, Mbbs MDDokumen59 halamanEmbryology of Cardiovascular System 1: DR - Navajyothi, Mbbs MDcheckmateBelum ada peringkat
- Lecture - 3 Cell Metabolism: Dr. S. Annie Jeyachristy Lecturer, Unit of Biochemistry Faculty of MedicineDokumen15 halamanLecture - 3 Cell Metabolism: Dr. S. Annie Jeyachristy Lecturer, Unit of Biochemistry Faculty of MedicinecheckmateBelum ada peringkat
- 7 - EnzymesDokumen15 halaman7 - EnzymescheckmateBelum ada peringkat
- BDS Cell StructureDokumen66 halamanBDS Cell StructurecheckmateBelum ada peringkat
- BDS-Batch 11-Module 1 - CA - Anatomy Question BankDokumen2 halamanBDS-Batch 11-Module 1 - CA - Anatomy Question BankcheckmateBelum ada peringkat
- 24 - Development of Jaw & PalateDokumen27 halaman24 - Development of Jaw & PalatecheckmateBelum ada peringkat
- 21 - Pharyngeal Arches (HD)Dokumen40 halaman21 - Pharyngeal Arches (HD)checkmateBelum ada peringkat
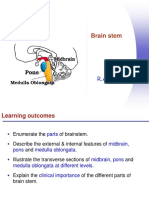
- 22 - Brain Stem (Neu)Dokumen64 halaman22 - Brain Stem (Neu)checkmate100% (3)
- Primary and Secondary MetabolitesDokumen27 halamanPrimary and Secondary Metabolitesshubham sabaleBelum ada peringkat
- Cellular Respiration ExplanationDokumen2 halamanCellular Respiration Explanationvestermail50% (2)
- Derived LipidsDokumen89 halamanDerived LipidsMichelleCurva67% (3)
- MT202 Unit5Dokumen52 halamanMT202 Unit5Nurul Balqish Abdul WahabBelum ada peringkat
- Tomato Jams Processing: Introduction/IdeasDokumen4 halamanTomato Jams Processing: Introduction/IdeasCandy Concepcion PadizBelum ada peringkat
- Chapter 8 QuizDokumen13 halamanChapter 8 QuizEbbie RoyaleBelum ada peringkat
- (Done) Biosynthesis of CholesterolDokumen25 halaman(Done) Biosynthesis of CholesterolFrancis Cedrick VictorinoBelum ada peringkat
- HECPDokumen35 halamanHECPaltaf ahmedBelum ada peringkat
- Module in Eng For Acad - First Quarter PDFDokumen53 halamanModule in Eng For Acad - First Quarter PDFJhien NethBelum ada peringkat
- Gluconeogenesis 2015Dokumen28 halamanGluconeogenesis 2015Secret AgentBelum ada peringkat
- Sulfur Slide ShowDokumen14 halamanSulfur Slide ShowAbdulRehmanBelum ada peringkat
- Hole's Essentials of Human Anatomy & Physiology Twelfth Edition - Chapter 4 Lecture OutlineDokumen47 halamanHole's Essentials of Human Anatomy & Physiology Twelfth Edition - Chapter 4 Lecture OutlineCharles AvilaBelum ada peringkat
- Cytoplasmic Labile Iron Accumulates in Aging Stem Cells Perturbing A Key Rheostat For Identity ControlDokumen71 halamanCytoplasmic Labile Iron Accumulates in Aging Stem Cells Perturbing A Key Rheostat For Identity ControlValen EstevezBelum ada peringkat
- Translation: Protein Biosynthesis Mrna Amino AcidsDokumen4 halamanTranslation: Protein Biosynthesis Mrna Amino AcidsMalick AwanBelum ada peringkat
- Unit 3 Biology WebquestDokumen5 halamanUnit 3 Biology WebquestEllie T. BrandBelum ada peringkat
- Introduction To Biochemistry CarbohydratesDokumen49 halamanIntroduction To Biochemistry Carbohydrateswanderer_1010Belum ada peringkat
- Regulation Cytoplasmic PH Bacteria: of inDokumen20 halamanRegulation Cytoplasmic PH Bacteria: of inEspacio UniversitarioBelum ada peringkat
- Enzymes of MitochondriaDokumen15 halamanEnzymes of MitochondriaHIRA JUNAIDBelum ada peringkat
- Cell Respiration Cornell Notes TeacherDokumen5 halamanCell Respiration Cornell Notes TeacherErik Artur100% (1)
- Nutrition in Plants - Mineral Nutrition: Module - 2Dokumen13 halamanNutrition in Plants - Mineral Nutrition: Module - 2keerthyBelum ada peringkat
- KetoGenic Diet PresentationDokumen33 halamanKetoGenic Diet PresentationAyat RajpootBelum ada peringkat
- Nutrition and Feed TechnologyDokumen29 halamanNutrition and Feed TechnologyAmitabh ChoudhuryBelum ada peringkat
- Price List - PT BIOMEDIKAL INDOPHARM NUSANTARADokumen2 halamanPrice List - PT BIOMEDIKAL INDOPHARM NUSANTARAhermanto soesilo100% (1)
- Industrial Wastewater Treatment by AS PDFDokumen404 halamanIndustrial Wastewater Treatment by AS PDFGustavo Mayen100% (1)
- Metabolisme GlikogenDokumen23 halamanMetabolisme GlikogenNurul AuliaBelum ada peringkat
- N CycleDokumen39 halamanN CyclePallabi DasBelum ada peringkat
- Adenosine TriphosphateDokumen2 halamanAdenosine TriphosphateThanh NguyenBelum ada peringkat
- Respiration BiologyDokumen20 halamanRespiration BiologySaidatul Atyah Mohd ApendaiBelum ada peringkat
- BecadexaminDokumen14 halamanBecadexaminHimanshu JhaBelum ada peringkat
- CCATMBTDokumen68 halamanCCATMBTAmit MandothiaBelum ada peringkat