Anda mungkin juga menyukai
- Práctica #4 - Morfología de La HojaDokumen9 halamanPráctica #4 - Morfología de La HojaLuis Miguel Arismendiz ImánBelum ada peringkat
- BIOELECTRICIDADDokumen19 halamanBIOELECTRICIDADsk9703Belum ada peringkat
- ALTIERI Principios EstrategiasDokumen8 halamanALTIERI Principios EstrategiasAndrés ChuraBelum ada peringkat
- Final 103 II 11 PatronDokumen3 halamanFinal 103 II 11 PatronemillyBelum ada peringkat
- Genes y Expresión GénicaDokumen64 halamanGenes y Expresión Génicaprimero militarBelum ada peringkat
- Establecimiento de La GestacionDokumen7 halamanEstablecimiento de La GestacionmarizhgBelum ada peringkat
- Investigación DocumentalDokumen10 halamanInvestigación DocumentalMariluz Arce SanchezBelum ada peringkat
- NeurotransmisoresDokumen14 halamanNeurotransmisoresJosué Reséndiz MendozaBelum ada peringkat
- Ficha PalomasDokumen1 halamanFicha PalomasLuis JimenezBelum ada peringkat
- Resumen Respectivo Del Tema 3 y 4Dokumen3 halamanResumen Respectivo Del Tema 3 y 4Yhamir Axel Apaza MamaniBelum ada peringkat
- Nutrientes de La Dieta y Apoptosis Como Mecanismos Reguladores Del CáncerDokumen8 halamanNutrientes de La Dieta y Apoptosis Como Mecanismos Reguladores Del CáncerJuan AspilcuetaBelum ada peringkat
- Contribución Al Conocimiento y Manejo Agroecologico de La Problematica Fitosanitaria Cardinal Del Cultivo de PlatanoDokumen68 halamanContribución Al Conocimiento y Manejo Agroecologico de La Problematica Fitosanitaria Cardinal Del Cultivo de PlatanoArmando NanotecBelum ada peringkat
- Ugaz Saavedra Johana - Prueba Molecular 20-01Dokumen1 halamanUgaz Saavedra Johana - Prueba Molecular 20-01Luna U100% (1)
- Ramsarsites Criteria SPDokumen2 halamanRamsarsites Criteria SPJean Berti Capcha RamosBelum ada peringkat
- Evolución biológica, especies y especiaciónDokumen2 halamanEvolución biológica, especies y especiaciónJenny Castro Jara100% (2)
- Viaje Al Interior Del Cuerpo HumanoDokumen5 halamanViaje Al Interior Del Cuerpo HumanojuanaBelum ada peringkat
- Resultado LaboratorioDokumen4 halamanResultado LaboratorioJose Alejandro Ramirez100% (1)
- Extracto de Semilla de Pomelo PDFDokumen44 halamanExtracto de Semilla de Pomelo PDFPurpaixão Produtos NaturaisBelum ada peringkat
- Mejoramiento en Cepas IndustrialesDokumen4 halamanMejoramiento en Cepas IndustrialesMariana NavarroBelum ada peringkat
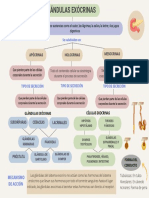
- Glándulas exocrinas: tipos y funcionesDokumen1 halamanGlándulas exocrinas: tipos y funcionesEsmeralda Camarillo OrtizBelum ada peringkat
- La Cadena Alimentaria Es Una RepresentaciónDokumen4 halamanLa Cadena Alimentaria Es Una RepresentaciónchabesabeBelum ada peringkat
- Glosario de BotanicaDokumen6 halamanGlosario de BotanicajudithdelgaBelum ada peringkat
- Complemento 2017Dokumen33 halamanComplemento 2017Criss SanchezBelum ada peringkat
- Feria335 01 Cultivo de Tejidos Clonacion Invitro de EpidendrumDokumen20 halamanFeria335 01 Cultivo de Tejidos Clonacion Invitro de EpidendrumSamoBelum ada peringkat
- Alteraciones morfológicas de los hematíesDokumen1 halamanAlteraciones morfológicas de los hematíesRogelio Velasquez100% (2)
- AlgasDokumen11 halamanAlgasEzequiel GuardiaBelum ada peringkat
- GastrulacionDokumen4 halamanGastrulacionElenasilvaLBelum ada peringkat
- Problemas para Resolver Encuentro 2 B L PracticaDokumen2 halamanProblemas para Resolver Encuentro 2 B L PracticaJonatan Caiza17% (6)
- Symbiosis Fungal-PlantDokumen11 halamanSymbiosis Fungal-PlantSantiago GuzmanBelum ada peringkat
- Ecodiversidad y Ecodiversidad en Honduras5Dokumen34 halamanEcodiversidad y Ecodiversidad en Honduras5Cecilia Banegas0% (1)