Anatomi Telinga
Diunggah oleh
pandu putraAnatomi Telinga
Diunggah oleh
pandu putra1
BAB I
PENDAHULUAN
Mendengar mempunyai peran penting sebagai sikap wapada terhadap
segala jenis aktivitas di sekitar kita, juga dalam mengapresiasi music, dan
khususnya palam proses komunikasi antar sesama. Suara melalui udara
menggetarkan tympanum, yang mana akan ditransmisikan ke tulang-tulang
pendengaran (maleus, inkus dan stapes) pada telinga tengah. Bergerak seperti
piston, Tulang pendengaran yang terakhir ini secara berurutan meningkatkan dan
menurunkan tekanan dalam koklea.2
Meski
pendengaran sangat dekat dengan kehidupan kita sehari-hari
namun jarang kita perhatikan keefektifannya.
Penampilan Telinga manusia
sungguh luarbiasa dalam beragam cara. Sensitivitas sangat tinggi sehingga kita
dapat mendeteksi sinyal yang menggetarkan gendang telinga sampai sekitar 1
picometer. Walaupun pendengaran kita meliputi frekuensi rentang 20 Hz sampai
20kHz, kita dapat membedakan sumber suara dan suasana bicara karena resolusi
frekuensi dapat memanjang sampai 1/30 dari interval antar iringan suara piano.5
Organ corti merupakan reseptor suara pada telinga manusia, sensori
epithelium berjalan sekitar 33 mm sepanjang spiral koklea. Komponen utama
organ corti adalah sel rambut, reseptor sensoris dari telinga dalam. Koklea sel
rambut ada 2 type, masing-masing mempunyai fungsi yang berbeda-beda dalam
proses transduksi. Dari 3.500 sel rambut dalam, yang tersusun dalam 1 baris,
membentuk sinaps dengan rata-rata 10 sel saraf afferent. Axon-axon saling
berhubungan dalam menyampaikan informasi pendengaran ke nucleus cochlearis
di otak. Meskipun 12.000 sel rambut luar dalam 3 baris juga dapat membuat
ikatan yang renggang pada saraf-saraf afferent, prinsipnya adalah mekanik : sel
rambut luar memperantarai proses aktif koklea.
Proses mendengar melingkupi beberapa tahap sebagai berikut. Suara
memasuki telinga, kemudian menggetarkan membrane tympani. Getaran ini
dihantarkan oleh rangkaian tulang pendengaran ke jendela oval. Disini, gerakan
getaran dari rangkaian tulang pendengaran ditransmisikan ke dalam cairan koklea,
yang mana akan menstimulasi reseptor sel rambut organ korti. Ketika sel rambut
berespon, sel-sel ini akan mengaktivasi saraf-saraf pada saraf pendengaran. Sinyal
ini kemudian dibentuk sebagai kode neural yang dapat diproses selanjutnya oleh
sistem saraf.
Telinga luar dan tengah secara berasama-sama disebut system konduktiv
karena sebagaian besar berfungsi menghantarkan sinyal suara dari udara ke dalam
telinga dalam. Koklea dan saraf cranial delapan menyusun sistem sensorineural,
disebut demikian karena meliputi respon fisiologi terhadap stimulus, aktivasi selsel saraf terkait dan penandaan respon sensori kedalam sinyal saraf. Aspek sistem
saraf pusat yang berhubungan dengan pengkodean pesan disebut sistem saraf
pusat auditori.
BAB II
ANATOMI TELINGA
2.1
Telinga Luar
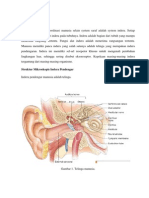
Telinga Luar terdiri atas Pinna (auricula) dan CAE ( Canalis Auricle
externa) dari meatus sampai membran tympani (gbr.1). Pinna manusia terbentuk
sebagian besar kartilago dan tidak ada otot. Bagian tengah dari pinna, concha,
merupakan bagian depan dari meatus auditori yang mana panjangnya sekitar 2,5
cm. Sepertiga lateral kanal merupakan kartilago. Bagian ini terdiri dari glandula
yang memproduksi serumen dan folikel rambut. Dua pertiga medial sisanya
merupakan tulang, termasuk didalamnya garis epitelial sampai membran
tympani.1 Kanalis auditorius eksterna berakhir pada membrane tympani. Kulit
dalam kanal mengandung kelenjar khusus, glandula seruminosa, yng mensekresi
substansi seperti lilin yang disebut serumen. Mekanisme pembersihan diri telinga
mendorong sel kulit tua dan dan serumen ke bagian luar telinga.3
Gambar 1. Telinga Luar1
Telinga luar dan dan kepala merupakan bagian yang pasif namun
mempunyai peran penting dalam pendengaran yang merupakan properti akustik.
Concha atau mangkuk auricula, mempunyai resonansi sekitar 5 kHz dan
permukaan ireguler dari pinna memperkenalkan resonance dan antiresonance.
Gambaran akustik berguna membantu membedakan apakah sumber suara di
depan atau belakang pendengar. 1
CAE merupakan tabung yang terbuka pada salah satu ujungnya dan
tertutup pada ujung lainnya, oleh karena itu CAE merupakan seperempat dari
gelombang resonansi. Frekuensi resonansi ditentukan oleh panjang dari tabung,
lengkungan tabung irelevan. Untuk tabung 2,5 cm, frekuensi resonansi sekitar 3,4
kHz.1
F0 : kecepatan suara @ 350 m/s/(4x2,5 cm)
Sifat akustik dari telinga luar adalah salah satu alasan gangguan
pendengaran akibat kebisingan pertama kali terjadi dan paling menonjol pada
wilayah frekuensi 4 kHz (boilermaker notch). Selain keunggulan noise-induced
hearing loss di wilayah 4 kHz, sifat akustik dari kepala dan telinga luar memiliki
peran penting dalam beberapa fungsi pendengaran . Di lokalisasi sumber suara,
kepala bertindak sebagai attenuator pada frekuensi di mana lebar kepala lebih
besar dari panjang gelombang suara. Demikian pada frekuensi lebih besar dari 2
kHz, terjadi head shadow effect, di mana perbedaan intensitas interaural dari 5
sampai 15 dB digunakan untuk melokalisasi sumber suara. Pada frekuensi rendah,
di mana panjang gelombang suara yang lebih besar dari lebar kepala, sedikit
pelemahan penangkapan gelombang dilakukan oleh kepala. Perbedaan waktu
Interaural (0.6 ms untuk suara dapat berjalan melintasi kepala) adalah syarat
penting untuk lokalisasi suara. head shadow effect adalah alasan pemburu
menggunakan senapan ditangan kanan memiliki gangguan pendengaran yang
lebih besar di telinga kiri daripada di telinga kanan mereka dan sebaliknya.
Moncong senapan, di mana energi akustik terbesar, lebih dekat ke telinga kiri dan
telinga kanan dilindungi oleh head shadow effect. 10 sampai 15dB gain yang
diberikan oleh telinga luar pada daerah 3 -5 kHz berguna untuk meningkatkan
deteksi dan pengenalan dari energi rendah, suara frekuensi tinggi seperti frikatif
tak bersuara. Pentingnya akustik sifat dari telinga luar dan kepala tercermin dalam
desain Alat Bantu Mendengar dan evaluasinya. Akhirnya, resonansi dari kanal
akustikus eksternal adalah pada bayi sekitar 8 kHz dan nilai menurun pada dewasa
setelah sekitar usia 2,5 tahun . Fitur perkembangan ini memiliki beberapa
implikasi klinis, terutama untuk pengujian sound field dan untuk desain alat bantu
mendengar dan evaluasi bayi1.
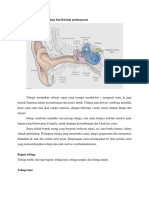
2.2
Telinga Tengah
Telinga tengah merupakan rongga berisi udara merupakan rumah bagi
osikuli (tulang telinga tengah) dihubungkan dengan nasofaring melalui tuba
eustachius, dan berhubungan dengan beberapa sel berisi udara di bagian mastoid
tulang temporal.3 Membran tympani merupakan dinding luar dari telinga tengah,
dengan bagian dalam berupa koklea. Batas atas dari tulang tengah membentuk
tulang diantara lobus otak tengah dan lantai otak tengah melapisi pembuluh darah
vena besar yang mengaliri darah dari kepala, bulbus jugularis. 3 Dua otot lurik,
yakni otot tensor tympani dan stapedius terletak di dalam telinga ini. Awalnya
menempel pada maleus dan dipersarafi. Otot stapedius melekat pada stapes dan
dipersarafi oleh cabang stapedial nervus facial.3
Satu fungsi dari otot telinga luar yaitu melindungi koklea dari suara keras, 1
Ketika suara keras yang melebihi 80 dB, terjadi reflex kontraksi dari otot
stapedius. Kontraksi ini meningkatkan kekakuan dari rangkaian tulang dan
membran tympani, menipiskan suara sampai kurang dari 2 kHz.
Suara dikonduksi dari membran tympani ke telinga dalam oleh tiga tulang
pendengaran yaitu maleus, inkus dan stapes. Maleus melekat pada belakang
membran tympani. Kepala maleus melekat pada dinding telinga tengah dan bagian
pendeknya melekat pada inkus yang pada akhirnya berartikulasi dengan kepala
stapes. Plat kaki pada stapes terpasang oleh ligamentum melingkar pada dinding
jendela oval, pembukaan ke dalam vestibuli telinga dalam atau koklea, yang mana
berartikulasi dengan tulang sendi stapedio-vestibular.3
Telinga tengah mentransmisikan energi akustik dari CAE yang berisi udara
sampai koklea yang berisi cairan. Fungsinya sebagai penyetara impedansi antara
udara yang berimpedansi rendah dengan impedansi tinggi dari koklea yang terisi
cairan. Kesesuaian impedansi ini dicapai dalam tiga langkah. Faktor pertama dan
yang terpenting adalah efektifitas area vibrasi membran tympani yaitu sekitar 1720 kali lebih besar dibanding efektifitas area vibrasi kaki tulang stapes (Gambar
2). Faktor kedua melingkupi aksi rangkaian tulang pendengaran. Lengan prosesus
incus lebih pendek, dengan faktor 1,3, dibandingkan dengan panjang manubrium
dan leher malleus. Ketiga dan merupakan faktor minor adalah bentuk membran
tympani. Hasil kombinasi ketiga faktor ini adalah pencapaian tekanan sekitar 2530 dB.1
Gambar 2. Skematik rangkaian tulang dan membran tympani menunjukkan
perbedaan di area dan pola vibrasi tulang-tulang1
Telinga tengah dengan tulang pendengarannya membentuk sistem
pengungkit untuk menghantarkan suara dari membran timpani ke fenestra ovale.
Transmisi energi suara melalui telinga tengah ke telinga dalam diawali dengan
membran timpani yang menggerakkan maleus. Membrana timpani menampung
getaran suara dari telinga luar, bekerja sebagai fibrator dan menjaga agar getaran
suara tidak langsung mengenai jendela bundar. Menurut Von Bekesy, pada
frekwensi sampai 2400 Hz membrana timpani bergerak menyerupai gerakan
piston, dimana pinggir atas membrana timpani dekat sulkus timpani sebagai
engsel sumbu putar dan pinggir bawah melakukan gerakan paling besar
mengelilingi sumbu putar. Pada frekwensi tinggi diatas 2400 Hz membran timpani
kehilangan kelenturannya sehingga gerakannya menjadi segmental.1,3
Getaran di gendang telinga kemudian ditransmisikan ke rangkaian tulang
pendengaran, yang mana getaran tersebut secara esential dalam bidang ke kanan
dan kiri. Getaran ini ditampilkan sebagai gerak berputar dari footplate stapes
dalam jendela oval Gambar 3)
Gambar 3. Telinga tengah mentransmisikan sinyal ke koklea melalui gerakan
berputar footplate stapedial pada jendela ovale.3
Otot tensor timpani dipersarafi nervus trigeminal ( saraf cranial N. V).
Yang terletak dalam semicanal tensor tympani di dinding superior anterior telinga
tengah dengan tuba eustachius. Kontraksi tensor timpani ini akan mendorong
maleus ke arah anteromedial, sehingga terjadi pengerasan rangkaian tulang
pendengaran.3
10
Otot stapedius merupakan otot skeletal tubuh terkecil,dengan panjang
sekitar 6,3 mm. Otot ini berada dalam eminence piramid dinding posterior cavum
timpani dan dipersarafi oleh nervus fascialis ( sarafa cranial N VII ). Kontraksi
stapedius mendorong stapes ke arah posterior. Meskipun kedua tulang ini terlihat
bergerak berlawanan namun keduanya mengeluarkan kekuatan yang tegak lurus
dengan gerakan normal tulang pendengaran.3
Aspek mendasar dari akustik (stapedius) reflex arc sebagai berikut. Proses
alur sensoris (aferen) melalui nervus auditori ke nukleus koklea ventral, dan
kemudian ke olivari superior komplex pada kedua sisi batang otak (menyilang sisi
berlawanan dari badan trapezoid). Alur motorik (eferen) mengikuti secara
bilateral, dan dari nukleus motorik nervus fascialis kedua sisi, kemudian melalui
nervus fasialis ke otot stapedius. Alur motorik aktivasi tensor timpani berjalan dari
nukleus nervus trigeminal ( nervus cranial V) ke otot tensor timpani melalui
nervus trigeminal. Karena kontraksi dari otot tensor timpani mengeraskan sistem
telinga tengah, transmisi frekuensi rendah berkurang.3
2.3
Telinga Dalam
2.3.1
Koklea
11
Telinga dalam berbentuk seperti pipa di dalam pipa. Pipa luar disebut osseus atau
tulang labyrinth karena dindingnya dibentuk diselubungi tulang. Pipa bagian
dalam terbentuk atas materil membrane dan kemudian disebut membrane
labyrinth (gambar 4).
Gambar 4. Penanda osseus (a) dan membranous (b) labyrinth3
Cochlea manusia berbentuk rumah siput, tabung tulang rata-rata
panjangnya 35 mm, terbagi atas skala vestibule, skala media dan skala tympani
(gbr 5). Skala vestibule dan tympani terdiri atas perilimfe, cairan extra selluler
12
seperti material dengan konsentrasi potassium (kalium) 4 mEq/L dan konsentrasi
natrium 139 mEq/L. Skala media terikat pada membrane basalis dan lamina
osseous spiral, dan dinding lateral. Skala ini terdiri atas endolimfe, cairan mirip
intrasel dengan kadar kalium 144 mEq/L dan natrium 13 mEq/L (gambar 6). Skala
media memiliki arus langsung positif (DC) potensial rest sekitar 80 mV yang
secara lambat menurun dari basis ke apex. Potensi endocochlear ini diproduksi
oleh pembuluh darah besar stria vaskularis dari dinding cochlea lateral. Pompa
Natrium-kalium-adenosin triphosphatase (Na+- K+-ATPase) dalam jumlah tertentu
sel-sel stria vaskularis berkontribusi terhadap potensi ini.
Gambar 5. Koklea terbagi atas skala vestibule, skala media dan skala tympani5
13
Gambar 6. Penampang midmodiolar ductus cochlea1
Energi akustik memasuki cochlea melalui semacam piston dari kaki-kaki
tulang stapes pada oval window dan tergandeng langsung terhadap perilimfe skala
vestibule. Perilimfe dari skala vestibuli berhubungan dengan perilimfe dari skala
tympani melalui lubang kecil di apex cochlea yang dikenal dengan helicotrema.1
jendela oval di dasar skala vestibule, dan jendela bulat di ujung basal skala
tympani. Skala media terpisah dari skala vestibule diatasnya oleh membrane
Reissners dan dari skala tympani dibawah oleh membrane basilar. Perlu
diperhatikan bahwa membrane basilaris paling sempit pada dasar dan secara
progresif melebar ke arah apex.3
Membran Labyrinth dari system pendengaran dan keseimbangan
terhubung oleh pipa tipis yang disebut ductus reunions, membentuk sistem
14
endolimfe kontinus. Sebagai tambahan akuaduktus koklearis berajalan dari
pembukaan skala tympani sampai ruang subarachnoid.3
Organ corti terletak pada membrane basalis dan tulang lamina spiral (gbr
7). Membran basilaris rata-rata lebar 0.12 mm pada basis dan meningkat rata-rata
0.5 mm pada apex. Komponen utama dari organ corti adalah rambut dalam dan
rambut luar, sel-sel penyangga (Deiters, Hensen, Claudius), membrane tektorial
dan plate lamina reticular kompleks (gbr 8). 3 Sel-sel penyokong membantu
struktur dan metabolik untuk organ corti. Proses phalang pada sel-sel Deiter dari
sel junctional dari lamina reticular.
Gambar 7. Organ Corti di atas membrane basalis5
15
Gambar 8. Gambaran sel rambut dalam 1 baris, dan sel rambut luar 3 baris3
Sel-sel rambut luar dan dalam organ corti sangat penting dalam transduksi
energi mekanik menjadi energi listrik. Sel-sel rambut luar sangat jauh berbeda dari
sel-sel rambut dalam. Gambar 9 dan tabel 1 secara detail terlihat perbedaannya.
Sebagai tambahan gambaran morfologi berbeda antara sel-sel rambut luar dan
dalam, demikian pula mengenai inervasi sarafnya. Ganglion spiral, badan sel dari
nervus auditorius, mengirimkan axon ke nucleus cochlea batang otak, dimana
projeksi dendrite melalui tulang lamina propia. Dari 50.000 saraf yang
menginervasi cochlea, 90% sampai 95% sinaps secara langsung pada sel-sel
rambut dalam, yang kemudian disebut neuron type I. Masing-masing sel rambut
dalam dipersarafi sekitar 15-20 neuron type I. Kebalikannya, 5% sampai 10% dari
50.000 saraf yang mempersarafi sel-sel rambut luar (neuron type II). Masingmasing neuron type II bercabang untuk mempersarafi sekitar 10 sel-sel rambut
luar. Sebagai tambahan dalam mempersarafi saraf afferent cochlea, sekitar 1800
16
serabut saraf eferen, bearasal dari ipsilateral dan kontralateral complex olivary
superior, yang memproyeksi kan ke cochlea.1
Gambar 9. Gambar skematik (a) sel rambut dalam dan (b) sel rambut luar. 3
17
Tabel 1. Struktur dan persarafan sel-sel rambut dalam dan luar1
Karakter
Jumlah
Bentuk
Sterosilia
Jumlah sel-sel rambut
Susunan
Sel-sel rambut dalam
3.500
Botol labu
Sel-sel rambut luar
12.000
Silindris
Sedikit
Banyak
Tiga atau empat baris, baris- Enam atau tujuh baris, basisbarisnya sedikit melengkung
Perlekatan
ke
membrane Koneksi
tidak
ada
barisnya tersusun bentuk V
atau W
atau Sterosilia terpanjang melekat
tektoria
Ultrastruktur
Posisi badan sel inti
Organel sitoplasms
renggang
kuat
Tengah
Terpencar
Basis
Bedekatan dengan membrane
Presynaptic khusus
Luas
sel
Kecil atau tidak ada (batang
Isi Glikogen
Hubungannya
dengan
Rendah
sel Lengkap disekitar
penyokong
Persarafan afferent
Sel ganglion
Tipe I
Jumlah sel-sel ganglion
27.000
Rasio se rambut terhadap 1.8:1
bersinaps dan vesikel)
Tinggi
Hanya
menyokong
di
permukaan dan basis
Tipe II
2.100
5.7:1
ganglion
Persarafan efferen
Sumber
Kompleks olivary superior Kompleks olivary superior
Target postsynaptic
lateral
Dendrite afferent
medial
Basisi sel rambut
Dari cochlea dapat direkam di CAE. Karena energi echo lebih besar daripada
sinyal durasi pendek, proses aktif, amplifikasi cochlea diperkirakan. Faktor yang
18
berkontribusi terhadap amplifikasi cochlea termasuk pergerakan sel-sel rambut
luar dan properti mekanik dari stereosilia dan membrane tektoria.
2.3.2
Proses aktif koklea
Pergerakan membrane basilar, ikatan rambut dan komponen osilasi lainnya
dari telinga berlawanan dengan tarikan hydrodinamika, yang mana menentukan
kekuatan terhadap kecepatan gerak. Viskositas merupakan lawan dari proses
mendengar yangmana suara cenderung akan tersebar. Pada koklea mammalia,
proses aktif sel rambut luar memperkeras getaran pada setiap kali penambahan
pada membrane basalis, menyokong energy hantaran gelombang.
Selama
gelombang berjalan, aktivitas beruntun sel rambut luar melawan efek dari
viskositas. Meskipun efek ini lemah pada posisi basal yang mana puncak
gelombang, kumulatif dari amplifikasi. Proses aktif dapat memperkuat oskilasi
lebih dari 1000 kali lipat. Untuk itu diperlukan penilaian yang seksama terhadap
suplai kekuatan proses aktif.4
Proses aktif koklea mammalian terdiri atas 2 komponen. Pertaman, organel
sel rambut yang sensitive secara mekanik, ikatan sel rambutnya dapat
menghasilkan kekuatan mekanik yang menambah gerakan membrane basilar.
Proses aktif mammalian yang kedua dikenal dengan nama motilitas somatic atau
electromotility.4
Komplek sel rambut stereosilia merupakan komponen penting untuk
transduksi. Stereosilia merupakan ikatan sekumpulan filament aktin yang
berbentuk tabung dan diinsersi ke dalam plate cuticular dan ada keterkaitan
19
diantaranya. Stereocilia sel rambut dalam memungkinkan tidak kontak dengan
membrane tektoria, tetapi pada sel rambut luar terdapat kontak langsung. Defleksi
stereocilia dengan riakan gelombang membuka dan menutup channel ion
nonspesifik pada ujung stereocilia, menyebabkan aliran kalium ke dalam sel
sensori. Aliran ion kalium ke dalam sel sensori dimodulasi dengan membuka dan
menutupnya channel ion stereocilia. Aliran potassium disebabkan oleh potensial
endocochlear +80 mV ditambah potensial negative sel-sel rambut. Akibat
depolarisasi intaselluler menyebabkan kaskade enzyme termasuk kalsium. Hal ini
mendorong pelepasan transmitter kimia, dan aktivasi serabut saraf aferen.1
Meskipun anggapan cochlea sebagai organ aktif dibanding organ pasif
tidak lagi menjadi perdebatan, detail spesifik dari amplifikasi cochlea dan basis
biologis masih terus diteliti. Satu hal yang melengkapi amplifikasi koklea
terhadap kemampuan sel rambut untuk berkontraksi dan memperpanjang dalam
proses sinyal elektrik, yang disebut somatic electromotility. Sebuah protein yang
bernama prestin telah diidentifikasi dalam sel rambut luar dan dianggap sebagai
penggerak protein sel-sel rambut luar dan memberikan kekuatan elektromotiliti sel
rambut. Sudut pandang lainnya memfokuskan pada aksi ion kalium dan kalsium
diduga menjadi basis amplifikasi koklea dan regulasi itu sendiri. Pendekatan yang
ke tiga mempertimbangkan bahwa kumpulan motor protein dalam sel rambut
dapat menggerakkan oksilasi yang tergantung pada elastisitas selnya.1
Neurotransmitter system saraf aferen dan eferen merupakan subjek yang
intens dipelajari. Glutamat merupakan neurotransmitter aferen. Glutamate telah
dideteksi pada sel ganglion spiral dan sel sensoris. Substansi transmitter utama
20
serabut saraf koklea adalah acetylcholine. Hal ini memungkinkan organ korti
modifikasi secara mekanik melalui perubahan motilitas sel-sel rambut dibawah
pengaruh sistem eferen. Aksi acetylcholin pada receptor menghasilkan
hiperpolarisasi dan melipatgandakan konduktans input sel. Reseptor acetylkolin
bersifat muskarinik dan nikotinik. Sebagai tambahan acetylkolin, asam
aminobtirat dan beberapa peptide neuroaktif merupakan neurotransmitter system
eferen.1
2.3.4
Potensial Gross koklea
Empat gross potensial (ekstraselluler) dapat terekam di koklea. Potensial
endolimfe (endokoklea), mikroponik koklea, potensial summating, dan potensi
aksi saraf umumnya. Tidake seperti potensial koklea lainnya, potensial
endolimfatik tidak digerakkan oleh stimuli respon akustik, namun, itu merupakan
potensial DC 80-100 mV terekam dalam skala media. Yang berjalan dari stria
vaskularis ke diding lateral dari koklea. Stria vaskularis dianggap sebagai sumber
energi, atau baterei koklea, yang mana sangat krusial untuk transduksi. Secara
alami sumber energi yaitu pompa Na+- K+-ATPase. Pompa ini telah dilokalisisr
untuk beberapa tipe sel koklea, termasuk sel marginal dari strai vaskularis, sel
sulkus luar dan fibrosit dekat dengan perlekatan membrane Reissner dan ligament
spiral. Sementara itu Na+-K+-ATPase harus berperan secara signifikan dalam
transport ion di koklea.1
21
Malfungsi dari mekanisme termasuk produksi endolimfe dan potensial
endolimfatik dapat mengakibatkan kehilangan pendengaran, yang disebut
metabolik presbiakusis. Ketika aliran endolimfe melalui duktus terhambat,
tekanan endolimfe akan meningkat dan terjadi kebocoran.1
Mikroponik koklea merupakan AC voltase terekam dalam koklea atau
dekat dengan jendela bulat. Hal ini memperlihatkan aliran ion melalui sel rambut
luar utamanya. Yang berarti bahwa resistens elektrik dari sel rambut luar diubah
oleh gerakan membrane basilar. Ketika stereosilia menekuk menjauh dari
modiolus, resistensi sel rambut menurun. Hasilnya adalah meningkatnya aliran
arus dan sedikit penurunan potensial endolimfatik. Ketika stereosilia menekuk
mendekati modiolus, resistensi meningkat dan aliran arus menurun seiring
meningkatnya potensial endolimfatik.
Kaitannya dengan fluktuasi voltase,
mikroponik koklea tergantung pada keberadaaan sel rambut luar. Tidak seperti
potensial neural, bentuk gelombang mikroponik koklea cermin dari gerakan
membrane basilar. Potensial akhir adalah potensial terekam pada koklea sebagai
respon terhadap suara.1,6
2.3.5
Fisiologi Nervus Delapan
Pada manusia nervus auditori mempunyai sekitar 30.000 serabut dan
50.000 pada kucing. Mungkin hal yang paliang penting peneliti temukan barubaru ini adalah 90% sampai 95% saraf (tipe I, serabut radial) yang mempersarafi
sel rambut dalam, sementara 5% sampai 10% (tipe II, serabut spiral luar yng
mempersarafi sel rambut luar (gambar 10). Paling banyak, jika tidak, serabut saraf
22
auditori berasal serabut terbesar tipe I yang berikatan dengan sel rambut dalam.
Serabut saraf radial ini mempunyai badan sel bipolar dalam ganglion spiral.
Serabut saraf spiral luar merupakan monopolar dan tidak bermyelin (gambar 11).1
Banyak dari serabut saraf mamalia gugur jika tidak ada stimulasi akustik.
Serabut saraf diklasifikasikan kedalam 3 kategori pada basis dengan rata-rata
gugur spontanTinggi (18-120 spike per detik), medium (0,5 18 spike per
detik), dan rendah (0 0,5 spike per detik).
23
Gambar 10. Hubungan antara sel rambut dalam, serabut radial dalam, dan
tipe I neuron auditori dalam ganglion spiral, dan sel rambut luar, serabut saraf
spiral, dan tipe II saraf auditori 3
Gambar 11. Gambaran skematik menunjukkan permukaan organ korti 1.
Distribusi serabut saraf koklea ke sel rambut dalam dan luar. Ujung basis koklea
menuju ke kanan. Banyak sinaps sel ganglion spiral dengan sel rambut dalam.
Masing-masing sel ganglion mempersarafi hanya satu sel rambut; beberapa sel
ganglion berikatan dengan tiap sel rambut. Hanya sekitar 5% sel ganglion spiral
mempersarafi sel rambut luar. Masing-masing serabut berjalan di dasar dalam
beberapa jarak sel rambut luar1
24
Serabut dengan rata-rata aktivitas spontan tinggi terhadap sinyal auditori
pada level rendah dibanding serabut dengan level medium maupun rendah ratarata akstivitas spontannya. Dengan kata lain, serabut dengan rata-rata spontanitas
tinggi memiliki dendrite yang tebal yang mana cenderung untuk keluar pada
bagian sisi rambut dalam terhadap sel rambut luar. serabut dengan rata-rata
spontanitas medium dan rendah memiliki dendrite yang tipis yang mana
cenderung terkikis pada bagian sisi rambut dalam terhadap modiolus.3
25
BAB III
FISIOLOGI PENDENGARAN
Rata-rata suara yang dapat di dengar manusia sekitar 16 - 20.000 Hz,
sensitivitas menurun pada frekuensi yang ekstrim, namun kemudain menjadi lebih
sensitive diatas 128 Hz sampai sekitar 4.000 Hz. Sensitivitas maximum dan
minimum berkurang sesuai umur.
Kepala itu sendiri berperan sebagai barrier alami antara dua telinga dan
kemudian sumber suara pada satu sisi akan menstimuli produksi lebih intens
terhadap telinga terdekat untuk kemudian akan sampai ke telinga tersebut dengan
cepat, yang mana hal ini dapat membantu dalam memberikan lokasi suara
berdasar pada intensitas dan waktu kedatangan suara yang berbeda-beda.
Frekuensi suara yang tinggi lebih penting dibanding frekuensi suara yang rendah,
hal ini menjelaskan kenapa lokasi suara menjadi lebih sulit pada tuli nada tinggi.4
Saluran telinga berperan sebagai pipa resonansi dan secara pasti
menguatkan
suara
antara
3000-4000
Hz
menambah
kecenderungan untuk rusak) telinga pada frekuensi ini.
sensitivitas
(dan
26
Telinga sangat sensitive dan berespon terhadap suara dengan intensitas
sangat rendah, untuk vibrasi yang mana sangat besar disbanding gerakan bebas
molekul udara. Untuk mencapai hal ini tekanan udara antar kedua membrane
tympani harus seimbang.4
Telinga luar dan telinga tengah berperan menguatkan sinyal suara. Pinna
dengan luas permukaan yang lebar dan corong mempresentasikan suara ke
membrane tympani yang kecil.; sebaliknya permukaan membrane tympani itu
sendiri lebih lebar dibanding footplate tulang stapes, sehingga ditemui adanya
penguatan hydraulic; gerakan kecil terhadap area yang lebar dikonversikan
sebagai gerakan yang kuat terhadap area yang kecil. Sebagai tambahan, rantai
tulang pendengaran merupakan penguat suara. Telinga luar dan telinga tengah
menguatakan suara dalam perjalanannya dari luar ke telingah dalam sekitar 30
dB.4,6
3.1
Telinga dalam
Fungsi dari telinga dalam adalah transduksi getaran ke dalam impuls saraf.
Sementara itu, hal ini juga menghasilkan analisis frekuensi (nada) dan intensitas
(keras) suara. Serabut saraf akan berjalan rata-rata 200 kali/detik. Level informasi
suara akan sampai di otak rata-rata 200 denyutan per detik.4
Seperti dijelaskan diatas, area sepanjang membrane basilar memiliki
karakteristik frekuensi sendiri-sendiri. Respon frekuensi tinggi pada basal
membrane dan respon frekuensi rendah pada bagia apex. Juga beberapa suara
diperkenalkan pada jendela oval oleh gerakan stapes yang ditransmisikan
27
sepanjang membrane basal sebagai gelombang hantar sampai semua komponen
frekuesi mencapai tempat repektif resonansi masing-masing.
Hanya sel rambut dalam yang menginisiasi impuls saraf yang terdengar
sebagai suara. Intinya dimana getaran membrane basalis yang terbanyak adalah
sekitar pertengahan sehingga sel rambut dalam bertahan sebagai getaran yang
kasar dari suara dengan intensitas yang tinggi. Pertanyaannya adalah, bagaiama
sel rambut dalam berespon terhadap sejumlah kecil setimulasi? Disini sel rambut
luar memiliki peran besar. Sel-sel ini akan terangsang oleh arus gelombang yang
berespon aktif dan kontraksi secara fisik. Mereka memiliki protein dalam dinding
mereka sendiri dan memendek. Karena kedua menempel pada membrane
reissners dan membrane basalis, menghasilkan gerakan tambahan pada
membrane labyrinth yang mana akan menguatkan arus gelombang pada
rangsangan maksimal.4
Jika sel rambut luar rusak, tidak ada lagi kontraksi dalam berespon
terhadap suara kecil dan sel rambut dalam juga tidak terstimulai. Hal ini akan
mengasilkan kehilangan pendengaran suara nada rendah.4
Stereosilia sel rambut sangat penting untuk proses mekanotransduksi.
Stereosilia adalah berkas serabut aktin yang membentuk pipa dan masuk ke dalam
lapisan kutikular. Membengkoknya stereosilia kearah stereosilia yang lebih tinggi
mengakibatkan terjadinya peregangan pada serabut tip link yang berada pada
puncak stereosilia yang mengakibatkan terjadinya pembukaan pintu ion pada
puncak stereosilia, menimbulkan aliran arus (K+) ke dalam sel sensoris. Aliran
28
kalium timbul karena terdapat perbedaan potensial endokohlea +80 mV dan
potensial intraselular negatif pada sel rambut, sel rambut dalam -45 mV dan sel
rambut luar -70 mV. Hal tersebut menghasilkan depolarisasi intraselular yang
menyebabkan kalium mengalir termasuk kalsium ke dalam sel rambut, kemudian
terjadi pelepasan transmiter kimia dari ruang presinaps yang berada pada dasar sel
rambut ke ruang sinaps dan akan ditangkap oleh reseptor serabut afferent n. VIII
menghasilkan potensial aksi yang akan diteruskan ke serabut n. VIII menuju
nukleus kohlearis.1
Pada saat kohlea mendapat stimulasi suara, maka akan terjadi perubahan
gerakan stereosilia yang diakibatkan terjadinya proses traveling wave pada
membrane basilaris yang mengakibatkan terjadinya pergerakan sel rambut kohlea
kearah stereosilia yang paling tinggi (depolarisasi) yang diikuti oleh terbukanya
ion channels serabut aktin pada puncak stereosilia sehingga terjadi Influx kalium
yang mengakibatkan terjadinya perubahan potensial intrasellular. Hal ini tercatat
sebagai cochlear microphonic dan summating potential. Kedua hal ini akan
tercatat pada berbagai bagian kohlea yang mempunyai frekuensi yang berbeda.
Potensial maksimum yang terjadi akan dicatat pada tiap frekuensi yang mencapai
titik maksimal amplitudo. cochlear microphonic dan summating potential ini
proses utamanya terjadi pada sel rambut luar. Stimulasi pergerakan stereosilia
pada sel rambut dalam sebagian besar dipengaruhi oleh pergerakan cairan
endolimf yang diakibatkan oleh pergerakan sel rambut luar, sedangkan pergerakan
membran basilaris mempunyai pengaruh yang lebih kecil terhadap pergerakan sel
rambut dalam. Hal inilah yang menjadi alasan kenapa cochlear microphonic pada
29
sel rambut luar dapat mencerminkan keadaan pada kohlea. Kerusakan pada sel
rambut luar kohlea secara total akan membuat penurunan pendengaran sekitar 60
dB, sehingga pada pemeriksaan OAE tidak akan memberikan respon.1
3.2
Proses Auditori Sentral
Impuls saraf dihantarkan oleh nervus VIII (nervus static-acoustic) dari
koklea sampai batang otak. Berikut serabut saraf yang mecapai nucleus dimana
mereka meneruskan serabut saraf lainnya. Serabut-serabut dari masing-masing
saraf auditori, beberapa mencapai sel sebrangnya sementara yang lainnya masih di
sisi yang sama,
3.2.1
Kemampuan untuk menyingkirkan suara yang tidak diinginkan
Di dalam ruangan yang berisik seorang pemuda dengan pendengaran
normal dapat mendengarkan atau tidak mendengarkan percakapan yang dia
kehendaki. Ini yang kita kenal degan teknik efek pesta koktail.
3.2.2
Lokasi ruang
Telinga manusia normal dapat melokalisir suara tenang
secara akurat.
Seseorang akan mengetahui dari arah mana datangnya suara; seseorang akan
menoleh mencari sumber suara, demikian juga dengan pesawat ataupun burungburung. Ada sel khusus yang berkaitan dengan hal seperti ini dengan otak
tengah.6,7
30
Penyilangan serabut saraf auditori terjadi pada beberapa level. Badan
trapezoid, lemniscus lateralis dan diantara coliculi lamina quadrigeminal. Diantara
pusat ini berlangsung proses inhibisi pada sisi dimana telinga distimulasi
kemudian oleh impuls dari telinga yang sebelumnya sudah dirangsang. Kemudian
nucleus koklea ini akan menginhibisi nucleus olivary superior, nucleus olivary
superior menginhibisi nucleus lemniscus lateral pada sisi sebelahnya. Nucleus
lemniscus latealis dapat menginhibisi pelepasan synaps colliculi inferior dari
lamina quadrigeminal dan colliculus itu sendiri menginhibisi badan geniculatum
medial pada sisi sebelahnya.6
Saraf pada lapisan bawah yang terstimulasi lebih dahulu menginhibisi
saraf pada lapisan yang lebih tinggi pada sisi yang bersebrangan (terstimulasi
kemudian) dan tidak diperkenankan untuk menginhibisi sisi yang terstimulasi
kemudian. Berikut sistem jalur yang menginhibisi satu sama lain (gbr 12 )6,12
31
Gambar 12. Auditori Pathway6
Anda mungkin juga menyukai
- Anatomi dan Fisiologi Telinga TengahBelum ada peringkatAnatomi dan Fisiologi Telinga Tengah22 halaman
- Proses Pendengaran dan Anatomi TelingaBelum ada peringkatProses Pendengaran dan Anatomi Telinga8 halaman
- Fisiologi Sistem Pendengaran ManusiaBelum ada peringkatFisiologi Sistem Pendengaran Manusia26 halaman
- Indra Pendengaran dan Reproduksi WanitaBelum ada peringkatIndra Pendengaran dan Reproduksi Wanita33 halaman
- Perambatan Bunyi Melalui Tulang TengkorakBelum ada peringkatPerambatan Bunyi Melalui Tulang Tengkorak22 halaman
- Proses Fisiologi Pendengaran ManusiaBelum ada peringkatProses Fisiologi Pendengaran Manusia15 halaman
- Laporan Kelompok Tutorial Ske 1 Blok THTBelum ada peringkatLaporan Kelompok Tutorial Ske 1 Blok THT45 halaman
- Evaluasi dan Rehabilitasi PendengaranBelum ada peringkatEvaluasi dan Rehabilitasi Pendengaran18 halaman
- Jurnal Fisiologi Blok SI 2 Yuda Monang S 219210042Belum ada peringkatJurnal Fisiologi Blok SI 2 Yuda Monang S 2192100426 halaman
- Tuli Mendadak: Diagnosis dan PenangananBelum ada peringkatTuli Mendadak: Diagnosis dan Penanganan21 halaman
- Sistem Pendengaran: Fisika Suara dan AnatomiBelum ada peringkatSistem Pendengaran: Fisika Suara dan Anatomi13 halaman
- Struktur dan Fungsi Sistem PendengaranBelum ada peringkatStruktur dan Fungsi Sistem Pendengaran25 halaman
- Anatomi dan Fisiologi Telinga ManusiaBelum ada peringkatAnatomi dan Fisiologi Telinga Manusia83 halaman
- Perambatan Bunyi Melalui Tulang TengkorakBelum ada peringkatPerambatan Bunyi Melalui Tulang Tengkorak13 halaman
- Fisiologi dan Anatomi Pendengaran ManusiaBelum ada peringkatFisiologi dan Anatomi Pendengaran Manusia59 halaman
- Anatomi Telinga Dan Fisiologi PendengaranBelum ada peringkatAnatomi Telinga Dan Fisiologi Pendengaran7 halaman
- Tatalaksana NIHL dan Hasil AudiometriBelum ada peringkatTatalaksana NIHL dan Hasil Audiometri21 halaman
- Fisiologi Telinga dan Pendengaran ManusiaBelum ada peringkatFisiologi Telinga dan Pendengaran Manusia14 halaman
- Fungsi dan Fisiologi Telinga ManusiaBelum ada peringkatFungsi dan Fisiologi Telinga Manusia6 halaman
- Uji Kepekaan Indera Pendengar ManusiaBelum ada peringkatUji Kepekaan Indera Pendengar Manusia21 halaman
- Struktur dan Mekanisme Telinga ManusiaBelum ada peringkatStruktur dan Mekanisme Telinga Manusia12 halaman
- Trauma Akustik: Penyebab dan PencegahanBelum ada peringkatTrauma Akustik: Penyebab dan Pencegahan26 halaman
- Penyakit Meniere: Gejala dan DiagnosisBelum ada peringkatPenyakit Meniere: Gejala dan Diagnosis35 halaman
- Anatomi dan Gangguan Telinga ManusiaBelum ada peringkatAnatomi dan Gangguan Telinga Manusia27 halaman
- Fisiologi Pendengaran dan Proses SuaraBelum ada peringkatFisiologi Pendengaran dan Proses Suara22 halaman
- Gangguan Pendengaran: Jenis dan PenyebabBelum ada peringkatGangguan Pendengaran: Jenis dan Penyebab41 halaman