Anda mungkin juga menyukai
- Model ABC TulipDokumen10 halamanModel ABC TulipTutik WulandariBelum ada peringkat
- Translated Copy of Turning Floral Organs Into LeavesDokumen17 halamanTranslated Copy of Turning Floral Organs Into LeavesERAPEWE OFFICIALBelum ada peringkat
- Abcde Model FloweringDokumen5 halamanAbcde Model FloweringErna grandeBelum ada peringkat
- Makalah Kuljar Bab 8Dokumen26 halamanMakalah Kuljar Bab 8Ricky RajuBelum ada peringkat
- PO 7 BIOLOGI - ALC Indonesia (WWW - Defantri.com)Dokumen15 halamanPO 7 BIOLOGI - ALC Indonesia (WWW - Defantri.com)elfira50% (2)
- Bab 5Dokumen30 halamanBab 5susan bureniBelum ada peringkat
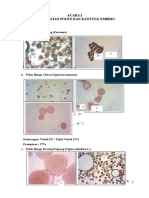
- Pengamatan Polen Dan Kantung Embrio'16Dokumen10 halamanPengamatan Polen Dan Kantung Embrio'16Dessy WulandariBelum ada peringkat
- LKP Botfar Morfologi 2021-FinalDokumen14 halamanLKP Botfar Morfologi 2021-Final206Arizal NurhakimF 21Belum ada peringkat
- Critical Book ReportDokumen3 halamanCritical Book ReportEwi Mellysa BarusBelum ada peringkat
- Soal Pat Biologi Kelas X Mia 2019-2020Dokumen15 halamanSoal Pat Biologi Kelas X Mia 2019-2020Siti NurmawatiBelum ada peringkat
- Jurnal Botani Jagung Versi IndoDokumen4 halamanJurnal Botani Jagung Versi IndoBem-dlm Faperta Unmer MadiunBelum ada peringkat
- Soal Us Bio Ix 2019Dokumen5 halamanSoal Us Bio Ix 2019sopianviaBelum ada peringkat
- Pertumbuhan Dan Perkembangan BungaDokumen52 halamanPertumbuhan Dan Perkembangan BungaRizki BestBelum ada peringkat
- Template TKA PlantaeDokumen6 halamanTemplate TKA PlantaeAhmad MujidinBelum ada peringkat
- Paper Evolusi Kelompok 2Dokumen4 halamanPaper Evolusi Kelompok 2MuhammadNasrulahAkbarBelum ada peringkat
- KrinyuDokumen10 halamanKrinyuMata ApiBelum ada peringkat
- Gen Identitas Organ (Organ Identity Gen) Dinda Mamangkay 17101102020Dokumen2 halamanGen Identitas Organ (Organ Identity Gen) Dinda Mamangkay 17101102020Alice WendyBelum ada peringkat
- ST BienertiaDokumen5 halamanST BienertiaJihan Sekar WijayantiBelum ada peringkat
- KELOMPOK 1 - Tugas Diksusi Praktikum BUNGADokumen32 halamanKELOMPOK 1 - Tugas Diksusi Praktikum BUNGANahidah AdillafarBelum ada peringkat
- Latihan Soal PersiapanBiologiDokumen41 halamanLatihan Soal PersiapanBiologiDanni SanjayaBelum ada peringkat
- Coretan PPT BRTDokumen5 halamanCoretan PPT BRTCelssy NataliBelum ada peringkat
- BAB II BarueeDokumen29 halamanBAB II BarueeLirofiatillahBelum ada peringkat
- Diskusi - Halaman 1-2Dokumen6 halamanDiskusi - Halaman 1-2Astrid ShabrinaBelum ada peringkat
- 2001 Soal Dan JawabanDokumen31 halaman2001 Soal Dan JawabanMuhamad JamilBelum ada peringkat
- Biologi Bunga Kembang Kertas Zinnia Elegans Jacq - Compress PDFDokumen12 halamanBiologi Bunga Kembang Kertas Zinnia Elegans Jacq - Compress PDFRifki AnnasBelum ada peringkat
- Muhammad Danur Ghiffari - B - K4321054 - Tugas 2 Riview JurnalDokumen3 halamanMuhammad Danur Ghiffari - B - K4321054 - Tugas 2 Riview JurnalMuhammad Danur GhiffariBelum ada peringkat
- Cabe Daun PDFDokumen10 halamanCabe Daun PDFZulfikasetiawanBudiBelum ada peringkat
- Struktur Anatomi Dan Perkembangan Pada Organ Bunga (Flos)Dokumen8 halamanStruktur Anatomi Dan Perkembangan Pada Organ Bunga (Flos)Muhammad SyahrullohBelum ada peringkat
- Tugas KuliahDokumen5 halamanTugas KuliahNur avina VinaBelum ada peringkat
- Kelompok 6 (Porang) - Makalah Ilmu Agronomi LanjutDokumen22 halamanKelompok 6 (Porang) - Makalah Ilmu Agronomi LanjutPriyo PrasetiyantoBelum ada peringkat
- Modul IDokumen8 halamanModul IResaadriannBelum ada peringkat
- 1.1 Naskah Soal Paket 1 - Mapel Biologi - Kelas X - Kurikulum 2013Dokumen11 halaman1.1 Naskah Soal Paket 1 - Mapel Biologi - Kelas X - Kurikulum 2013Agus SuhendraBelum ada peringkat
- LKM - Diagram - Bunga (1) PuputDokumen7 halamanLKM - Diagram - Bunga (1) PuputZidni FilhaqBelum ada peringkat
- Mekanisme Perkembangan Stomata Pandangan EvolusionerDokumen3 halamanMekanisme Perkembangan Stomata Pandangan EvolusionerFatima Niskala Intan PramanikBelum ada peringkat
- Biologi X IpsDokumen8 halamanBiologi X IpsEqi BaihaqiBelum ada peringkat
- Pengaruh Konsentrasi Ekstrak TaogeDokumen20 halamanPengaruh Konsentrasi Ekstrak TaogeEklips C' KhBelum ada peringkat
- Soal TODokumen2 halamanSoal TOAsniar AnggraeniBelum ada peringkat
- Soal PAT BIOLOGI KLEAS X KerjaDokumen12 halamanSoal PAT BIOLOGI KLEAS X KerjaEdenia KudoBelum ada peringkat
- Morfologi PADIDokumen21 halamanMorfologi PADIalohayahooBelum ada peringkat
- Keanekaragaman Tumbuhan Lumut Dan EvolusiDokumen53 halamanKeanekaragaman Tumbuhan Lumut Dan Evolusidwi rulitasari0% (2)
- 1Dokumen21 halaman1viriyacsgoBelum ada peringkat
- TumbuhanDokumen3 halamanTumbuhanNur Fahmi UtamiBelum ada peringkat
- Laporan Praktikum Bunga IndvDokumen14 halamanLaporan Praktikum Bunga Indveka nur wahyuniBelum ada peringkat
- Shoot Apical Meristem and Root Apical MeristemDokumen8 halamanShoot Apical Meristem and Root Apical MeristemDewi Ekaputri Pitorini100% (1)
- Sejarah GiberelinDokumen8 halamanSejarah Giberelinemiraa9679Belum ada peringkat
- Biologi UsbnDokumen68 halamanBiologi UsbnRizki Eka PutriBelum ada peringkat
- Quiz Bab 1Dokumen18 halamanQuiz Bab 1Anonymous dqfQczsdZn0% (2)
- Ipi 419251Dokumen4 halamanIpi 419251Indri LestariBelum ada peringkat
- Farmakognosi A1.1Dokumen23 halamanFarmakognosi A1.1Afifatul JannahBelum ada peringkat
- Template Pengamatan Polen Dan Kantung EmbrioDokumen9 halamanTemplate Pengamatan Polen Dan Kantung EmbrioYudhi RizaldiBelum ada peringkat
- Hormon ABC Pada PembungaanDokumen3 halamanHormon ABC Pada PembungaanAfan's BenWadd100% (1)
- PlantaeDokumen20 halamanPlantaeDirgahBelum ada peringkat
- LAPORAN Ke 8 PRAKTIKUM BIOLOGIDokumen4 halamanLAPORAN Ke 8 PRAKTIKUM BIOLOGIRisky bima PurnawanBelum ada peringkat
- File Soal Ujian Kelas Xii Sma N TaekasDokumen14 halamanFile Soal Ujian Kelas Xii Sma N TaekasRichard RanBelum ada peringkat
- Soal Anfistum Dan AnfiswanDokumen14 halamanSoal Anfistum Dan AnfiswanAdiSoekariawan100% (1)
- Analisa Soal Kelas XiDokumen303 halamanAnalisa Soal Kelas XiNugroho Widi SasmitoBelum ada peringkat
- 1 s2.0 S0254629916312807 MainDokumen5 halaman1 s2.0 S0254629916312807 MainEkha KristianaBelum ada peringkat