Metabolisme Energi
Diunggah oleh
Ranhii'siiepoetriforget MaaharaniiyHak Cipta
Format Tersedia
Bagikan dokumen Ini
Apakah menurut Anda dokumen ini bermanfaat?
Apakah konten ini tidak pantas?
Laporkan Dokumen IniHak Cipta:
Format Tersedia
Metabolisme Energi
Diunggah oleh
Ranhii'siiepoetriforget MaaharaniiyHak Cipta:
Format Tersedia
BAB.
VI
BIOENERGETIK NUTRISI
TUJUAN INSTRUKSIONAL UMUM
Pada akhir pokok bahasan ini pembaca diharapkan mampu memahami pengertian energi,
metabolisme karbohidrat, lemak, dan protein pada ikan
TUJUAN INSTRUKSIONAL KHUSUS
Pada akhir pokok bahasan ini pembaca diharapkan mampu :
1. Menjelaskan pengertian energi dan tingkatan energi biologis
2. Menjelaskan mekanisme metabolisme energi dari karbohidrat
3. Menjelaskan mekanisme metabolisme energi dari lemak
4. Menjelaskan mekanisme metabolisme energi dari protein
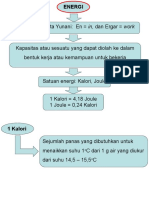
6.1. Pengertian Energi
Ikan membutuhkan energi untuk pertumbuhan, aktivitas dan reproduksi. Penggunaan
energi dalam tubuh ikan dijabarkan dalam bioenergetik. Istilah energi berasal dari Yunani,
yang terdiri dari kata "en" berarti di dalam dan "ergon" berarti kerja, sehingga energi dapat
didefinisikan sebagai suatu kemampuan untuk melakukan pekerjaan dan berbagai bentuk
kegiatan (kimia, elektrik, radiasi dan termal) dan dapat diubah. Energi radiasi dari matahari
yang digunakan tanaman untuk membentuk zat-zat makanan dapat digunakan oleh ternak
untuk menghasilkan kerja mekanik. Sebagian besar energi yang terdapat di permukaan bumi
berasal dari matahari, sedang energi yang digunakan untuk kerja adalah energi kimia yang
disimpan dalam pakan ikan. Energi dalam pakan umumnya disebut dengan energi biologis.
Energi biologis terdiri dari beberapa tingkatan sebagaimana terlihat pada Gambar 6.1.
Energi intake (IE) atau energi kotor (GE) adalah sejumlah panas yang dilepaskan oleh
satu unit bobot bahan kering pakan bila dioksidasi sempurna. Energi kotor bahan pakan
ditentukan dengan jalan membakar contoh bahan pakan dalam bom kalorimeter. Kandungan
IE biasanya dinyatakan dalam satuan Mkal IE/kg BK. Tidak semua IE bahan pakan dapat
dicerna, sebagian akan dikeluarkan bersama feses. Energi kotor dalam feses disebut sebagai
fecal energy. Energi feses ini selain berasal dari pakan yang tidak dicerna juga berasal dari
saluran pencernaan yang berupa mukosa, enzim dan bakteri.
Energi yang masuk tubuh/intake energy (IE) dari pakan
Energi feses/fecal energy (FE) yang hilang dari :
a. pakan yang tidak dicerna
b. produk metabolis (mukosa, bakteri dan enzim)
Energy tercerna/digestible energy (DE) yang meliputi juga :
a. Energi dari fermentasi saluran pencernaan
Energi urin/urinary energy (UE) yang hilang dari :
a. sisa hasil metabolisme
b. katabolisme endogenous
Energi insang/gills energy (ZE) energy yang hilang melalui insang
Energi termetabolis/metabolizable energy (ME) meliputi juga :
a. panas dari fermentasi pencernaan
Energi kenaikan produksi panas tubuh/heat increament energy (HE)
yang hilang terdiri dari :
a. Produksi panas pada pakan/the heat increament of feeding
(HiE)
b. Produksi panas akibat maintenance (HEm)
Energi bersih/nett energy (NEp + NEm)
Energi untuk hidup pokok/maintenance energy (NEm) :
a. Metabolisme basal
b. Aktivitas hidup pokok
c. Menjaga temperatur tubuh
Energi untuk tumbuh dan reproduksi/ recovered energy
(RE)
a. Disimpan dalam jaringan : tumbuh, lemak,
b. Disimpan dalam produk : telur dan fetus
c. Kerja
Gambar 6.1. Bagan energi
Pakan ikan umumnya menghasilkan energi feses sebasar 10 40% dari energi kotor
(energi yang dikonsumsi). Apabila data kecernaan energi dan nutrien pada ikan diketahui,
maka total energi feses dapat dihitung dengan rumus:
Ef = (100% - Ed) x Ec
Dimana :
Ef = energi fese
Ed = kecernaan energi
Ec = energi bruto
Energi tercerna (DE) adalah berapa banyak IE yang dapat dicerna dengan cara
mengurangi IE bahan pakan dengan GE feses. Satuan DE adalah Mkal DE/kg BK. Tidak
semua energi yang dicerna akan diserap. Energi termetabolis (ME) adalah energi kotor dari
pakan yang dapat digunakan oleh tubuh. Sebagian besar atau bahkan seluruh energi yang
dapat dimetabolisasi akan digunakan untuk proses metabolisme. Kebutuhan energi untuk
metabolisme harus dipenuhi dahulu dan apabila berlebih dapat digunakan untuk
pertumbuhan. Rumus energi termetabolsime adalah :
ME = Ei (EVf + EVu)
ME = metabolizable energi
Ei = energi yang dikonsumsi
Ef = energi feses
Eu = energi terbuang lewat ekskresi nitrogen
Produksi panas adalah (H) adalah energi yang berupa kenaikan produksi panas yang
terjadi akibat proses metabolisme dan fermentasi dari zat-zat makanan. Sampai dengan
pengukuran ME, pengukuran dengan teknik bom kalorimeter dapat digunakan. Pengukuran
HE tidak dapat lagi menggunakan bom kalorimeter, namun dengan teknik kalorimetri hewan.
Kenaikan produksi panas ini sebagian besar berasal dari metabolisme zat-zat makanan dalam
tubuh.
Energi termetabolis digunakan untuk aktivitas/pemeliharaan/hidup pokok
(maintenance) atau untuk pemeliharaan/hidup pokok beserta produksi. Secara umum energi
untuk pemeliharaan/hidup pokok disebut NEm dan energi untuk tumbuh dan bereproduksi
disebut RE. Ikan membutuhkan energi secara kontinyu untyuk maintenance tanpa melihat
apakah ikan mengkonsumsi pakan atau tidak. Ikan yang sedang dipuasakan akan
memperoleh energi dari cadangan energi tubuh. Energi maintenance digunakan untuk
metabolisme basal, dan menyokong tubuh pada saat istirahat. Metabolisme basal adalah
tingkat pembelanjaan energi minimal untuk mempertahankan struktur dan fungsi jaringan
tubuh agar hewan tetap survive.
NEm dalam tubuh digunakan untuk tetap dalam kondisi keseimbangan. Dalam
tingkat ini tidak terjadi penambahan atau pengurangan energi dalam jaringan tubuh. Nilai
NEm umumnya ditentukan dengan mengukur produksi panas ikan percobaan yang berstatus
gizi baik, dipuasakan, ada dalam lingkungan termonetral dan beristirahat. Produksi panas
ikan yang berada dalam kondisi seperti itu disebut "Basal Metabolic Rate". RE digunakan
untuk kerja diluar kemauan, pertambahan bobot jaringan (pertumbuhan, atau produksi
lemak), telur dan sebagainya.
Energi aktivitas pada ikan adalah energi untuk aktivitas berenang. Energi yang
dikeluarkan untuk berenang dapat mengurangi porsi energi yang sebenarnya dapat digunakan
untuk menyusun jaringan tubuh yang baru. Energi berenang tersebut dapat melebihi jumlah
energi yang dikonsumsi dan untuk keseimbangannya maka energi dipasok dari cadangan
energi.
Dari berbagai ketentuan diatas diartikan bahwa semua energi yang terdapat dalam
feses dan dalam urin dianggap hanya berasal dari pakan saja, dengan demikian maka nilai
DE, ME dan NE bukan merupakan nilai energi yang sebenarnya, akan tetapi merupakan nilai
energi semu atau nilai yang tampak atau apparent energy. Oleh karena itu untuk nilai energi
yang sebenarnya atau true energy harus dikoreksi terlebih dahulu dengan energi yang berasal
dari bukan sisa pakan atau yang disebut energi endogenous. Pengukuran ME pada ikan baik
dalam bentuk Apparent Metabolizabel Energy (AME) maupun True Metabolizable Energy
(TME) dapat dilihat pada Bab Evaluasi Pakan. Energi umumnya diperoleh dari zat-zat
makanan karbohidrat, lemak dan protein. Lemak menyediakan energi dua kali lipat per gram
mol dibandingkan dengan karbohidra dan protein. Setiap gram mol lemak akan
menghasilkan energi sebesar sekitar 9 kkal dibadingkan dengan 4 kkal pada karbohidrat dan
protein.
6.2. Metabolisme energi dari karbohidrat
Metabolisme karbohidrat untuk menghasilkan energi dimulai dari masuknya glukosa
asal darah ke dalam sel. Disini terjadilah proses glikolisis tahap pertama yang dimulai
dengan reaksi antara glukosa dengan ATP (adenosin tri phosphat) dengan adanya enzim
glukokinase (yang memerlukan ion Mg2
+
sebagai kofaktor) dalam rangka melakukan
fosforilasi (pemasukan satu gugus fosfat) glukosa menjadi glukosa-6-fosfat, dengan
menghasilkan ADP (adenosin di phosphat).
Reaksi tahap kedua merupakan isomerisasi glukosa-6-fosfat diubah menjadi fruktosa-
6-fosfat, yang dikatalisis oleh fosfoheksoisomerase. Dalam reaksi ini tidak terjadi penguraian
maupun pembentukan ATP. Reaksi tahap ketiga adalah pemasukan gugus fosfat dari ATP,
dikatalisis oleh fosfofruktokinase dengan ion Mg
2+
sebagai kofaktor dan terbentuklah
fruktosa-1,6-difosfat dengan meninggalkan lagi ADP.
Reaksi tahap keempat merupakan pemecahan senyawa karbohidrat beratom enam
menjadi dua senyawa beratom tiga. Fruktosa-1,6-difosfat dengan bantuan enzim aldolase,
dipecah menjadi dua molekul triosa fosfat yaitu 3, gliseraldehida 3-fosfat dan
dihidroksiaseton fosfat. Selanjutnya terjadi reaksi isomerisasi bolak-balik antara kedua
senyawa beratom tiga ini dikatalisis oleh triosafosfat isomerase. Dalam keadaan normal
dihidroksiaseton fosfat diubah seluruhnya menjadi gliseraldehida 3-fosfat sehingga
kemungkinan hilangnya setengah dari energi molekul glukosa dapat dicegah. Dapat
dikatakan disini, pemecahan satu molekul fruktosa 1,6-fosfat menghasilkan dua molekul
gliseraldehida 3-fosfat. Tahap-tahap reaksi satu sampai empat memerlukan energi dan gugus
fosfat dari penguraian ATP menjadi ADP.
Reaksi tahap kelima merupakan perubahan gliseraldehida 3-fosfat menjadi asam 1,3-
difosfogliserat, yang melibatkan reaksi pemasukan satu gugus fosfat dari asam fosfat (buka
dari ATP) dan oksidasi molekul aldehida menghasilkan molekul asam karboksilat. Reaksi ini
dikatalisis oleh gliseraldehida 3-fosfat dehidrogenase dan dirangkaikan dengan reaksi reduksi
pembentukan NADH (bentuk reduksi dari nikotinamid adenin dinukleotida) dari NAD
+
(bentuk oksidasinya). Reaksi tahap kelima dalam tahap glikolisis merupakan reaksi pertama
yang menghasilkan energi.
Tahap keenam, satu dari dua buah ikatan antara asam fosfat dengan asam gliserat
dalam molekul asam 1,3-difosfogliserat adalah suatu ikatan anhidrida yang dalam proses
pemecahannya menghasilkan energi untuk pembentukan ATP dari ADP dan Pi. Reaksi ini
dikatalisis oleh fosfogliserat kinase (dengan ion magnesium sebagai kofaktor) dengan
menghasilkan asam 3-fosfogliserat.
Reaksi tahap ketujuh adalah isomerasi asam gliserat 3-fosfat menjadi asam gliserat 2-
fosfat, dikatalisis oleh fosfogliserat mutase dengan ion magnesium atau ion mangan sebagai
kofaktor. Reaksi tahap kedelapan adalah enzim enolase melepaskan satu molekul H
2
O dari
asam gliserat 2-fosfat menghasilkan asam fosfoenolpiruvat dengan ion magnesium atau ion
mangan sebagai kofaktor.
Reaksi tahap kesembilan atau terakhir dari glikolisis adalah pembentukan asam
piruvat dari asam fosfoenolpiruvat melalui senyawa antara asam enolpiruvat. Dalam reaksi
yang dikatalisis oleh piruvat kinase (ion magnesium atau sebagai kofaktor) gugus fosfat
yang dilepaskan oleh fosfoenolpiruvat dipakai untuk mensintesis ATP dari ADP. Perubahan
enolpiruvat ke asam piruvat terjadi secara spontan.
Tahapan glikolisis secara menyeluruh dibagi menjadi dua bagian. Bagian pertama
meliputi tahap reaksi enzim yang memerlukan ATP, yaitu tahap reaksi dari glukosa sampai
dengan pembentukan fruktosa 6-fosfat, yang menggunakan dua molekul ATP untuk tiap satu
molekul glukosa yang dioksidasi. Bagian kedua meliputi tahap reaksi yang menghasilkan
energi (ATP dan NADH), yaitu dari gliseraldehida 3-fosfat sampai dengan piruvat. Dari
bagian kedua ini dihasilkan dua molekul NADH dan empat molekul ATP untuk tiap molekul
glukosa yang dioksidasi (atau untuk dua molekul gliseraldehida 3-fosfat yang dioksidasi).
Karena satu molekul NADH yang masuk rantai pengangkutan elektron dapat menghasilkan
tiga molekul ATP, maka tahap reaksi bagian kedua ini menghasilkan 10 molekul ATP.
Dengan demikian keseluruhan proses glikolisis menghasilkan 10 - 2 = 8 molekul ATP untuk
tiap molekul glukosa yang dioksidasi. Secara keseluruhan tahap glikolisis dapat dilihat pada
Gambar 6.2.
Selanjutnya asam piruvat diubah melalui salah satu jalur berikut ini.
1. Dapat masuk ke mitokondria lalu ikut dalam siklus asam trikarboksilat (siklus asam sitrat,
siklus Krebs) untuk melakukan oksidasi dan fosforilasi ADP menjadi ATP dalam sistem
sitokrom (ini adalah jalur yang paling sering terjadi pada asam piruvat).
2. Dapat direduksi membentuk asam laktat dan bersifat reversibel.
3. Dapat diubah kembali menjadi karbohidrat melalui glikoneogenesis (kebalikan dari
glikolisis).
4. Dapat direduksi kembali menjadi asam malat lalu masuk dalam siklus Krebs.
5. Dapat dioksidasi menjadi asam oksaloasetat dalam siklus Krebs.
6. Dapat diubah menjadi asam amino alanin melalui transaminasi.
Hal ini semua adalah jalur yang mungkin dijalani oleh asam piruvat, dan ini
tergantung pada metabolisme sel waktu itu. Selama proses glikolisis, setiap molekul glukosa
membentuk dua molekul asam piruvat yang kesemuanya terjadi di sito plasma sel.
Reaksi oksidasi piruvat hasil glikolisis menjadi atetil koenzim A merupakan tahap
reaksi penghubung yang penting antara glikolisis dengan jalur metabolisme lingkar asam
trikarboksilat (siklus Krebs). Reaksi yang dikatalisis oleh kompleks piruvat dehidrogenase
dalam matriks mitokondria melibatkan tiga macam enzim (piruvat dehidrogenase,
dihidrolipoil transasetilase dan dihidrolipoil dehidrogenase), lima macam koenzim 9-
tiaminpirofosfat, asam lipoat, koenzim A, flavin adenin dinukleotida dan nikotinamid adenin
dinukleotida), dan berlangsung dalam lima tahap reaksi.
Glikogen
Uridin difosfat glukosa
Glukosa Glukosa-1-P
Glukosa-6-P
Fruktosa-6-P
Fruktosa-1,6-diP
Gliseraldehida-3-P dihidroksi fosfat
1,3-di-P-gliserat
3-P-gliserat
2-P-gliserat
Fosfoenol piruvat
Melalui mitokondria
piruvat
Gambar 6.2. Tahapan glikolisis
Piruvat + NAD
+
+ koenzim A asetilkoenzim A + NADH + CO
2
Tahap reaksi pertama dikatalisis oleh piruvat dehidrogenase yang menggunakan
tiamin pirofosfat sebagai koenzimnya. Dekarboksilasi piruvat menghasilkan senyawa -
hidroksietil yang terikat pada gugus cincin tiazol dari tiamin pirofosfat. Pada tahap reaksi
kedua, -hidroksietil didehidrogenase menjadi asetil yang kemudian dipindahkan dari tiamin
pirofosfat ke atom S dari koenzim yang berikutnya, yaitu asam lipoat, yang terikat pada
enzim dihidrolipoil transasetilase. Dalam hal ini gugus disulfida dari asam lipoat diubah
menjadi bentuk reduksinya, yaitu gugus sulfhidril. Pada tahap reaksi ketiga, gugus asetil
dipindahkan dengan perantaraan enzimdari gugus lipoil pada asam dihidrolipoat, ke gugus
tiol (sulfhidril pada koenzim A). Kemudian asetilkoenzim A dibebaskan dari sistem enzim
kompleks piruvat dehidrogenase. Pada tahap reaksi keempat, gugus ditiol pada gugus lipoil
yang terikat pada dihidrolipoil transasetilase dioksidasi kembali menjadi bentuk disulfidanya
dengan enzim dihidrolipoil dehidrogenase yang berikatan dengan FAD (flavin adenin
dinukleotida). Pada tahap kelima atau terakhir, FADH
2
(bentuk reduksi dari FAD) yang
tetap terikat pada enzim, dioksidasi kembali oleh NAD
+
(nikotinamid adenin dinukleotida)
menjadi FAD, sedangkan NAD
+
berubah menjadi NADH (bentuk reduksi dari NAD
+
).
Siklus Krebs terjadi di dalam mitokondria dan membutuhkan oksigen agar dapat
berlangsung. Asam piruvat yang berasal dari glikolisis, begitu masuk ke dalam mitokondria
diubah menjadi asetil koenzim A. Kemudian bersamaan dengan berlangsungnya proses
oksidasi dalam siklus Krebs, pasangan-pasangan atom hidrogen (2H) dilepaskan bersama
dengan CO
2
. Atom-atom hidrogen tersebut menyajikan ion H
+
atau proton dan elektron
yang kemudian masuk ke dalam sistem transport elektron mitokondria. Ion hidrogen dan
elektron di pungut oleh molekul NAD
+
(nikotinamid adenin dinukleotid), mereduksi NAD
+
menjadi NADH. NADH merupakan pengantara siklus Krebs dan enzim dalam membran
dalam mitokondria yang akan mengangkut elektron melalui sistem sitokrom dari rantai
respirasi.
NADH mentransfer proton dan elektron dan terbentuklah FMN (flavin
mononukleotid). Kemudian menurut teori kemiosmotik, MFN mengambil proton dari bagian
dalam membran, hingga tereduksi menjadi FMNH
2
. Kemudian dua atom H-nya dilepaskan
dan ditransfer ke membran mitokondria eksterior dan dilepas berupa proton (H
+
). Pada saat
yang sama, dua elektron itu menggabung ke molekul ubikuinon atau koenzim Q, yang
kemudian mengambil atom-atom H. Kemudian dilepaskanlah satu elektron ke sitokrom C
1
dan lainnya ke sitokrom b dari membran mitokondria. Elektron-elektron kemudia ditransfer
ke sitokrom a dan a
3
, dari sinilah elektron bergabung dengan atom oksigen dan dua proton
untuk membentuk molekul air.
Dalam urutan oksidasi reduksi yang terjadi di dalam membran serta melintas
membran mitokondria, tiap dua proton yang melintas membran dan masuk, akan
menyebabkan fosfat anorganik melekat pada ADP karena adanya perbedaan potensial listrik,
lalu terbentuklah ATP. Kecepatan reaksi ini akan meningkat oleh adanya sistem enzim.
Hasil neto dari siklus Krebs serta sistem transport sitokrom adalah untuk
menghasilkan tiga ATP lebih banyak dari ADP untuk tiap pasang atom H yang dilepaskan
selama siklus tersebut, dan hal ini terjadi melalui fosforilasi oksidatif. Di sini juga dihasilkan
tiga molekul CO
2
dan tiga molekul H
2
O.
Karena ada dua molekul piruvat yang terbentuk dari tiap molekul glukosa, siklus
Krebs bekerja dua kali untuk tiap molekul glukosa yang dipecahkan. Oleh karena itu, pada
dasarnya akan diperoeleh empat pasang atom hidrogen untuk tiap siklus. Dua siklus akan
menghasilkan 8 x 3 = 24 ATP, dan dua ATP neto dari glikolisis, ditambah empat ATP lagi
dari pembentuk FAD yang tereduksi selama siklus Krebs. Di samping itu juga dua lagi ATP
dari fosforilasi oksidatif pada tingkat substrat, yang kesemuanya menjadi 32 ATP, enam lagi
masih mungkin dari generasi glikolitik dari NADH
2
.
Jadi dapat dinyatakan 38 molekul ATP dihasilkan dari degradasi satu molekul
glukosa. ATP yang terbentuk itu merupakan sumber energi yang siap untuk tiap kegiatan
biologi termasuk kontraksi otot, sekresi kelenjar, konduksi saraf, absorpsi aktif dan transport
membran. Secara keseluruhan siklus Krebs dapat dilihat pada Gambar 6.3.
Asetil KoA
KoA-SH
H
2
O
NADH + H
+
H
2
O
NAD
+
H
2
O
H
2
O
CO
2
FADH
+
NAD
+
NADH + H
+
FAD CO
2
+ 2H
H
2
O GDP + P
i
P
I
GTP
Gambar 6.3. Siklus Krebs
Piruvat, dengan adanya NADH, H
+
dan enzim laktik dehidrogenase, membentuk
laktat dan NAD. Dengan pengubahan yang bersifat enzimatis, laktat kemudian
dikonversikan kembali menjadi piruvat yang kemudian masuk siklus Krebs untuk oksidasi
lengkap seperti yang telah dikemukakan sebelumnya. Hasil akhirnya selalu CO
2
, H
2
O dan
energi yang siap digunakan dalam bentuk ATP. Secara keseluruhan metabolisme karbohidrat
dapat dilihat pada Gambar 6.4.
Sebagian dari glukosa yang masuk ke dalam sel tidak mengalami katabolisme menjadi
piruvat oleh glikolisis, tetapi membentuk glikogen secara anabolis melalui proses yang
disebut glikoneogenesis, sehingga glukosa untuk sementara dapat disimpan dalam hati.
Sitrat
Isositrat
Cis-aconitat
-ketoglutarat
Suksinil KoA
Suksinat
Fumarat
Malat
Oksaloasetat
SIKLUS
KREBS
NAD
+
NADH + H
+
Proses ini kemudian diikuti oleh proses kebalikannya, yaitu glikogenolisis yang merupakan
pemecahan cadangan glikogen menjadi glukosa-6-fosfat pada beberapa sel, atau langsung
menjadi glukosa seperti yang terjadi di hati.
Glukosa tidaklah harus selalu masuk ke sel dari kapiler darah. Beberapa sel terutama
sel hati, dapat menghasilkan glukosa dari substrat dan bukan dari karbohidrat. Hal ini adalah
pembentukan glukosa dari sel-sel lemak atau protein di dalam hati, untuk aliran darah, yang
disebut dengan proses glukoneogenesis. Hal ini pada dasarnya ini terjadi ketika tingkat
glukosa darah menurun, atau ketika jumlah glukosa yang masuk ke dalam sel tidak
mencukupi dan cadangan glikogen terpakai habis.
Gambar 6.4. Metabolisme karbohidrat
6.3. Metabolisme energi dari lemak
Asam palmitat (C16:0) merupakan salah satu asam lemak yang paling banyak
diketahui proses metabolismenya, oleh karena itu untuk memudahkan pembahasan
selanjutnya akan dipakai asam lemak ini. Proses penguraian asam lemak dimulai dengan
tahap -oksidasi. Proses oksidasi ini berlangsung dalam mitokondria. Tahap pertama adalah
Asetil KoA
KoA-SH
H
2
O
NADH + H
+
H
2
O
NAD
+
H
2
O
H
2
O
CO
2
FADH
+
NAD
+
NADH + H
+
FAD
CO2
CO
2
+ 2H
H O
GDP + P
i
P
i
GTP
Sitrat
Isositrat
Cis-aconitat
-ketoglutarat
Suksinil KoA
Suksinat
Fumarat
Malat
Oksaloasetat
SIKLUS
KREBS
NAD
+
NADH + H
+
H
2
O
Glikogen
Uridin difosfat glukosa
Glukosa
Glukosa-1-P
Glukosa-6-P
Fruktosa-6-P
Fruktosa-1,6-diP
Gliseraldehida-3-P dihidroksi fosfat
1,3-di-P-gliserat
3-P-gliserat
2-P-gliserat
Fosfoenol piruvat
piruvat
menggiatkan asam palmitat bebas dengan asetil koenzim A dalam sitoplasma, oleh enzim asil
koenzim A sintetase menghasilkan palmitoil koenzim A. Pada reaksi ini sebagai sumber
energi digunakan satu molekul ATP untuk satu molekul palmitil koenzim A yang terbentuk.
Dalam hal ini terjadi dua reaksi pemecahan ikatan fosfat berenergi tinggi, yaitu
terhidrolisisnya ATP menjadi AMP + PP
i
dan terurainya PP
i
menjadi 2 P
i
oleh enzim
pirofosfattase. Dengan demikian untuk menggiatkan satu molekul asam lemak dalam tahap
reaksi ini, digunakan energi yang didapatkan dari pemecahan dua ikatan fosfat berenergi
tinggi dari satu molekul ATP. Tahap reaksi kedua, palmitoil koenzim A diangkut dari
sitoplasma ke dalam mitokondria dengan bantuan molekul pembawa yaitu karnitin yang
terdapat dalam membran mitokondria.
Reaksi tahap ketiga adalah proses dehidrogenasi palmitoil koenzim A yang telah
berada di dalam mitokondria dengan enzim asil koenzim A dehidrogenase yang
menghasilkan senyawa enoil koenzim A. Pada reaksi ini FAD (flavin adenin dinukleotida)
yang bertindak sebagai koenzim direduksi menjadi FADH
2
. Dengan mekanisme fosforilasi
bersifat oksidasi melalui rantai pernafasan suatu molekul FADH
2
dapat menghasilkan dua
molekul ATP.
Pada tahap reaksi keempat, ikatan rangkap pada enoil koenzim A dihidratasi menjadi
3-hidroksipalmitoil koenzim A hidratase.
Reaksi tahap kelima adalah dehidrogenase dengan enzim 3-hidroksianil koenzim A
dehidrogenase dan NAD+ sebagai koenzimnya. Pada reaksi ini 3-hidroksipalmitoil koenzim
A dioksidasi menjadi 3-ketopalmitoil koenzim A, sedangkan NADH yang terbentuk dari
NAD+ dapat dioksidasi kembali melalui mekanisme fosforilasi bersifat oksidasi yang
dirangkaikan dengan rantai pernafasan menghasilkan tiga molekul ATP.
Reaksi tahap terakhir adalah mekanisme oksidasi- adalah pemecahan molekul
dengan enzim asetil koenzim A asetiltransferase atau disebut juga tiolase. Pada reaksi ini
satu molekul koenzim A (CoA) bebas berinteraksi dengan 3-ketopalmitoil keenzim A
menghasilkan satu molekul asetil koenzim A dan sisa rantai asam lemak dalam bentuk
koenzim A-nya, yang mempunyai rantai dua atom karbon lebih pendek dari palmitoil
koenzim A semula.
Proses degradasi asam lemak selanjutnya adalah pengulangan mekanisme oksidasi-
secara kontinu sampai rantai panjang asam lemak tersebut habis dipecah menjadi molekul
asetil koenzim A. Dengan demikian satu molekul asam palmitat (C16) menghasilkan 8
molekul asetil koenzim A (C2) dengan melalui tujuh kali oksidasi-. Setelah semua reaksi
-oksidasi berakhir maka dilanjutkan dengan masuk dalam siklus Krebs. Reaksi keseluruhan
dari katabolisme asam lemak palmitat dapat dilihat pada Gambar 6.5.
Biosintesis asam lemak dari asetil koenzim A terjadi di hampir semua bagian tubuh
ikan, terutama dalam jaringan hati dan jaringan lemak. Biosintesis ini berlangsung melalui
mekanisme yang dalam beberapa hal berbeda dengan oksidasi asam lemak. Secara
keseluruhan biosintesis asam lemak terbagi menjadi tiga tahap utama. Tahap pertama
pembentukan malonil koenzim A dari asetil koenzim A. Tahap kedua adalah pemanjangan
rantai asam lemak sampai terbentuknya asam palmitat secara kontinu dengan tiap kali
penambahan malonil keenzim A dan pelepasan CO
2
. Tahap ketiga adalah pemanjangan rantai
asam palmitat secara bertahap bergantung pada keadaan dan komposisi faktor penunjang
reaksi dalam sel.
Tahap pertama dimulai dengan reaksi antara asetil koenzim A dengan gugus SH
(sulfhidril) dari molekul ACP (acyl carrier protein) merupakan reaksi pemul dalam
mekanisme biosintesisi asam lemak. Reaksi ini dikatalisis oleh salah satu dari enam enzim
sintetase kompleks, ACP-asiltransferase, dengan persamaan reaksi :
Gambar 6.5. Katabolisme asam palmitat
Asetil-S-CoA + ACP-SH asetil-S-ACP + CoA-SH
Reaksi selanjutnya adalah pemindahan gugus asetil dari ACP ke gugus SH dari enzim
beta-ketoasil-ACP-sintase, menghasilkan asetil S-beta-ketoasil-ACP-sintase, disingkat asetil-
S-sintase.
Asetil-S-ACP + sintase-SH ACP-SH + asetil-S-sintase
ATP AMP
CoASH
PP
i
(1)
Asam lemak asil asam lemak koenzim
A FAD
(2)
FADH
Enoil koenzim
A H
2
O
Hidroksiasil koenzsim A
NAD
(4)
NADH + H
+
Ketoasil koenzim A
CoASH
(5)
Tiokinase
Palmitoil-CoA dehidrogenase
Enoil hidrase
Hidroksisiasil dehidrogenase
Tiolase
(3)
Asetil KoA
KoA-SH
H
2
O
NADH + H
+
H
2
O
NAD
+
H
2
O
H
2
O
CO
2
FADH
+
NAD
+
NADH + H
+
FAD
CO2
CO
2
+ 2H
H O
GDP + P
i
P
i
GTP
Sitrat
Isositrat
Cis-aconitat
-ketoglutarat
Suksinil KoA
Suksinat
Fumarat
Malat
Oksaloasetat
SIKLUS
KREBS
NAD
+
NADH + H
+
H
2
O
Dengan telah terikatnya gugus asetil pada enzim pertama dari enam enzim kompleks
sintetase asam lemak tersebut, dapatlah dimulai mekanisme pemanjangan rantai asam lemak
dengan penambahan dua atom karbon pada malonil koenzim , secara berturut-turut sampai
terbentuknya asam palmitat.
Tahap kedua adalah reaksi kondensasi pembentukan aseasetil-S-AC. Reaksi
kondensasi didahului dengan reaksi pembentukan malonil-S-ACP dari malonil-S-CoA, yaitu
pemindahan gugus malonil dari ACP ke CoA. Reaksi ini dikatalisis oleh enzim ACP-
malonil-transferase :
Malonil-S-CoA + ACP-SH malonil-S-ACP + CoA-SH
(malonil koenzim A) (koenzim A)
Reaksi berikutnya adalah kondensasi antara asetil-S-sintase dengan malonil-S-ACP
menghasilkan asetoasetil-S-ACP. Reaksi ini dikatalisis oleh enzim beta-ketoasil-ACP-sintase
dan laju reaksinya didorong oleh terlepasnya CO
2
dari malonil-S-ACP, yaitu reaksi
eksergonik dekarboksilasi gugus malonil, yang memberikan dorongan termodinamik ke arah
pembentukan aseto-asetil-S-ACP.
Pada tahap ketiga ini, terdapat dua reaksi reduksi asetoasetil-S-ACP. Pada reaksi
reduksi yang pertama, aseoasetil-S-ACP diredukis dengan NADPH dan enzim beta-ketoasil-
ACP-reduktase menghasilkan D--hidroksibutiril-S-ACP, yang selanjutnya mengalami
dehidratasi dengan enzim enoil-ACP-hidratase menghasilkan krotonil-ACP. Reaksi reduksi
yang kedua adalah hidrogenasi krotonil-ACP dengan enzim enoil-ACP-reduktase yang
menghasilkan butiril-ACP. Seperti juga reaksi reduksi yang pertama, reaksi ini
menggunakan NADPH-NADP
+
(bukan NADH-NAD
+
seperti yang dipakai pada proses
oksidasi asam lemak) sebagai sistem koenzimnya .
Dengan terbentuknya butiril-ACP, selesailah satu dari tujuh daur yang dilakukan oleh
enzim kompleks sintetase untuk menghasilkan palmitoil-CoA. Untuk memulai daur yang
berikutnya, gugus butiril dipindahkan dari ACP ke enzim -ketoasil-ACP-sintase dan ACP
mengambil satu gugus malonil dari molekul malonil Co-A yang lainnya. Selanjutnya daur
diulangi dengan reaksi kondensasi antara malonil-ACP dengan butiril-S--ketoasil-ACP
sintase menghasilkan -ketoheksanoil-S-ACP dan CO
2
. Demikianlah setelah tujuh kali
mekanisme daur berlangsung dengan enzim kompleks sintetase asam lemak, terbentuklah
palmitoil-ACP sebagai hasil akhir. Selanjutnya gugus palmitoil ini dapat mengalami
beberapa kemungkinan, tergantung kondisi dalam sel dan jenis jasadnya. Kemungkinan itu
adalah, pertama, gugus palmitoil dilepaskan dari enzim sintetase kompleks, dengan bantuan
enzim tioesterase, menghasilkan asam palmitat bebas, kedua, gugus palmitoil dipindahkan
dari ACP ke CoA, ketiga, gugus palmitoil digabungkan langsung ke dalam asam fosfatidat
dalam proses biosintesis fosfolipid dan triasil gliserol. Gambar 6.6 menunjukkan mekanisme
reaksi keseluruhan proses biosintesis asam palmitat dari asetil-CoA. Reaksi oksidasi dan
biosintesis asam palmitat mempunyai perbedaan yang cukup penting. Perbedaan tersebut
terdapat pada Tabel 6.1.
Tabel 6.1. Perbedaan oksidasi dengan biosintesis asam palmitat
No. Komponen Oksidasi Biosintesis
1. Tempat Mitokondria Sitoplasma
2. Sistem pembawa CoA ACP
3. Molekul pemanjang rantai Asetil-CoA (beratom karbon
dua)
Malonil-CoA (beratom karbon
tiga)
4. Sistem koenzim dalam
reaksi hidrogenasi
NAD
+
/NADH & FAD/FADH
2
NADPH/NADP
+
CO
2
CH
3
CO SCoA HOOCCH
2
CO SCoA
ACP-SH asetil-CoA malonil-CoA ACP-SH
CoASH CoASH
CH
3
CO S ACP HOOCCH
2
CO S ACP
Sintase-SH
ACP-SH
CH
3
CO S sintase
Sintase-SH
O COOH CO
2
CH
3
C CH CO S ACP CH
3
COCH
2
CO S ACP
Asetoasetil-S-ACP
NADPH + H
+
NADP
+
OH O
CH
3
C CH
2
C S ACP
-hidroksibutiril-S-ACP
H
2
O
CH
3
CH CH
2
CO S ACP CH
3
(CH
2
)
13
CH
2
COOH
NADPH + H
+
Krotonil-ACP asam palmitat
CoA-SH
NADP
+
CH
3
CH
2
CH
2
CO S ACP CH
3
(CH
2
)
13
CH
2
CO SCoA
Butiril-ACP Palmitoil-CoA
Reaksi diulangi mulai dari CoA-SH
tahap reaksi dengan sintase-SH
sampai dengan reaksi enoil-ACP reduktase
Diulangi sampai 6 kali ACP-SH
CH
3
(CH
2
)
13
CH
2
CO S ACP
Palmitoil-ACP
Gambar 6.6. Mekanisme reaksi keseluruhan proses biosintesis asam
palmitat dari asetil-CoA
ACP-asiltransferase
ACP-malonil
transferase
-ketoasil-ACP reduktase
enoill-ACP hidratase
enoill-ACP reduktase
6.4. Metabolisme energi dari protein
Metabolisme protein tidak secara langsung terlibat dalam memproduksi energi.
Tetapi metabolisme protein terlibat dalam produksi enzim, hormon, komponen struktural, dan
protein darah dari sel-sel badan dan jaringan. Metabolisme energi yang berasal dari protein
didahului dengan degradasi protein menjadi asam-asam amino. Kemudian asam-asam amino
dilepas gugus aminonya melalui deaminasi oksidatif di sel-sel hati. Hasil deaminasi akan
masuk dalam siklus Krebs guna pembentukan energi, atau melalui piruvat dan asetil koenzim
A sebelum masuk siklus Krebs.
Kerangka karbon dari asam-asam amino alanin, sistein, sistin, glisin, treonin, serin
dan hidroksiprolin diubah menjadi piruvat. Pembentukan piruvat dari glisin dapat terjadi
dengan konversi menjadi serin yang dikatalisis oleh enzim serin hidroksimetiltransferase.
Reaksi alanin transaminase dan serin dehidratase, keduanya memerlukan piridoksal
fosfat sebagai koenzim. Reaksi serin dehidratase berlangsung melalui pembuangan H
2
O dari
serin, membentuk suatu asam amino tidak jenuh. Kemudian disusun kembali menjadi asam
-amino yang terhidrolisis spontan menjadi piruvat dan amonia.
Jalan katabolik utama dari sistin adalah konversi menjadi sistein yang dikatalisis oleh
enzim sistin reduktase. Setelah itu akan bergabung dengan katabolisme sistein.
Sistein dikatabolisme melalui dua jalan katabolisme utama yaitu jalan oksidasi
langsung (sistein sulfinat) dan jalan transaminasi (3-merkaptopiruvat). Kedua jalan tersebut
memerlukan enzim transaminase.
Treonin dibelah menjadi asetaldehida dan glisin oleh treonin aldolase. Kemudian
asetaldehida membentuk asetil koenzim A, sementara glisin sudah dibicarakan diatas.
Tiga dari lima karbon 4-hidroksi-L-prolin dikonversi menjadi piruvat, dua sisanya
membentuk glikosilat. Kemudian tahap akhir reaksi melibatkan aldolase yang memecah
hidroksiprolin menjadi piruvat dan glioksilat.
Semua asam amino yang membentuk piruvat dapat dikonversi menjadi asetil koenzim
A. Disamping itu ada lima asam amino yang membentuk asetil koenzim A tanpa membentuk
piruvat lebih dahulu. Asam-asam amino tersebut adalah fenilalanin, tirosin, triptofan, lisin
dan leusin. Fenilalanin mula-mula dikonversi menjadi tirosin oleh fenilalanin hidroksilase.
Lima reaksi enzimatik berurutan mengkonversi tirosin menjadi fumarat dan asetoasetat, yaitu
(1) transaminasi menjadi p-hidroksifenilpiruvat, (2) oksidasi dan migrasi sekaligus dari rantai
samping 3-karbon dan dekarboksilasi yang membentuk homogentisat, (3) oksidasi
homogentisat menjadi maleilasetoasetat, (4) isomerasi maleiasetoasetat menjadi
fumarilasetofumarat dan (5) hidrolisis fumarilasetoasetat menjadi fumarat dan osetoasetat.
Asetoasetat selanjutnya dapat mengalami pembelahan tiolitik menjadi asetat dan asetil
koenzim A.
L-lisin dikonversi menjadi -aminoadipat dan -ketoadipat. L-lisin pertama kali
berkondensasi dengan -ketoglutarat yang memecah air dan membentuk basa Schiff.
Kemudian direduksi menjadi sakaropin oleh dehidrogenase dan kemudian dioksidasi oleh
dehidrogenase kedua. Penambahan air membentuk L-glutamat dan L--aminoadipat--
semialdehida. Katabolisme lebih lanjut dari -aminoadipat memerlukan transaminasi
menjadi -ketoadipat, yang mungkin diikuti oleh dekarboksilasi oksidatif menjadi glutaril-
KoA.
Triptofan oksigenase (triptofan pirolase) mengkatalisis pembelahan cincin dengan
inkorporasi 2 atom oksigen yang membentuk N-formilkinurenin. Oksigenasenya adalah
metaloprotein besiforfirin. Pengeluaran gugus formil dari N-formilkinurenin secara hidrolitik
dikatalisis oleh kinurenin formilase yang menghasilkan kinurenin. Kemudian dideaminasi
dengan transaminase gugus amino rantai samping ke ketoglutarat. Metabolisme lebih lanjut
dari kinurenin melibatkan konversi menjadi 3-hidroksikinurenin. Kinurenin dan
hidroksikinurenin dikonversi menjadi hidroksiantranilat oleh enzim kiruneninase suatu
enzim piridoksal fosfat. Leusin sebelum diubah menjadi asetil koenzim A diubah dahulu
menjadi asetoasetat, sama dengan pengubahan tirosin.
Suksinil koenzim A merupakan hasil akhir amfibolik dari katabolisme metionin,
isoleusin dan valin yang hanya sebagian rangka dikonversi. Empat per lima karbon valin,
tiga per lima karbon metionin dan setengah karbon isoleusin membentuk suksinil koenzim A.
l-metionin berkondensasi dengan ATP membentuk S-adenosilmetionin atau "metionin aktif".
Pengeluaran gugus metil membentuk S-adenosil-homosistein. Hidrolisis ikatan S-Peserta
menghasilkan L-homosistein dan adenosin. Homosistein selanjutnya berkondensasi dengan
serin, membentuk sistationin. Pembelahan hidrolitik sistationin membentuk L-homoserin dan
sistein. Kedua reaksi ini oleh karenanya juga terlibat dalam biosintesis sistein dan serin.
Homoserin dikonversi menjadi -ketobutirat oleh homoserin deaminase. Konversi -
ketobutirat menjadi propionil-KoA selanjutnya terjadi dengan cara biasa untuk dekarboksilasi
oksidatif asam -keto membentuk derivat asil KoA.
Sebagaimana diharapkan dari kemiripan strukturnya, katabolisme L-valin dan L-
isoleusin pada awalnya memerlukan reaksi yang sama. Jalan ini kemudian memisah dan
masing-masing rangka asam amino mengikuti jalan unik menjadi zat antara amfibolik.
Kerangka karbon dari asam-asam amino glutamin, glutamat, arginin, histidin, dan
prolin memasuki siklus Krebs melalui -ketoglutarat. Katabolisme glutamin dan glutamat
berlangsung dengan bantuan enzim glutaminase dan transaminase.
Prolin dioksidasi menjadi dehidroprolin yang dengan penambahan air akan
membentuk glutamat -semialdehida. Selanjutnya dioksidasi menjagi glutamat dan
ditransaminasi menjadi -ketoglutarat. Arginin dan histidin juga membentuk -ketoglutarat,
satu karbon dan baik 2 (histidin) maupun 3 (arginin) pertama-tama harus dikeluarkan dari
asam amino 6 karbon ini. Arginin hanya membutuhkan hanya satu langkah yaitu
pengeluaran gugus guanidino secara hidrolisis yang dikatalisis oleh arginase yang
menghasilkan ornitin. Ornitin mengalami transaminasi gugus -amino, membentuk glutamat
-semialdehida, yang dikonversi menjadi -ketoglutarat.
Bagi histidin, pengeluaran karbon dan nitrogen yang berlebih membutuhkan empat
reaksi. Deaminasi histidin menghasilkan urokanat. Konversi urokanat menjadi 4-
imidazolon-5-propionat, yang dikatalisis oleh urokanase melibatkan penambahan H
2
O dan
oksidasi-reduksi interna. Reaksi selanjutnya adalah 4-imidazolon-5-propionat dihidrolisis
menjadi N-formiminoglutamat yang diikuti oleh pemindahan gugus formimino pada karbon
alfa ke tetrahidrofolat yang membentuk N
5
-formiminotetrahidrofolat. Kemudian dengan
bantuan enzim glutamat formimino transferase, N
5
-formiminotetrahidrofolat diubah menjadi
L-glutamat dan akhirnya menjadi oksaloasetat dengan bantuan enzim transaminase.
Secara umum katabolisme masing-masing asam amino yang digunakan sebagai
sumber energi dapat dilihat pada Gambar 6.7.
Sebagaimana diketahui, sumber energi diperoleh dari karbohidrat, lemak dan protein.
Karbohidrat melalui jalur glikolisis dan kemudian menuju siklus Krebs untuk menghasilkan
energi. Lemak melalui jalur -oksidasi dan kemudian menuju siklus Krebs untuk
menghasilkan energi. Sementara itu protein harus mengalami deaminasi sebelum menjadi
piruvat, asetil koenzim A ataupun langsung masuk ke siklus Krebs. Kesemua jalur tersebut
dapat dilihat pada Gambar 6.8.
Amonia merupakan produk sisa utama dari katabolisme protein dapa ikan, sehingga
tidak dibutuhkan energi untuk memekatkannya dan menawarkannya seperti pada hewan
lainnya.
Alanin Glutamat Arginin
Triptofan Glutamin
Sistin Glutamat
Glisin Histidin
Treonin -ketoglutarat Prolin
Serin
Hidroksiprolin
Sitrat Suksinil KoA
Piruvat
Asetil KoA
Oksaloasetat Fumarat
Metionin
Isoleusin
Valin
Isoleusin
Leusin
Triptofan Asetoasetil KoA Aspartat Tirosin
Fenilalanin
Fenilalanin
Tirosin
Triptofan Asparagin
Lisin
Leusin
Gambar 6.7. Katabolisme asam-asam amino menjadi energi
Siklus Krebs
Karbohidrat Lemak Protein
Glukosa Asam lemak Asam amino
Gambar 6.8. Metabolisme energi
Asetil KoA
KoA-SH
H
2
O
NADH + H
+
H
2
O
NAD
+
H
2
O
H
2
O
CO
2
FADH
+
NAD
+
NADH + H
+
FAD
CO2
CO
2
+ 2H
H O
GDP + P
i
P
i
GTP
Sitrat
Isositrat
Cis-aconitat
-ketoglutarat
Suksinil KoA
Suksinat
Fumarat
Malat
Oksaloasetat
SIKLUS
KREBS
NAD
+
NADH + H
+
H
2
O
Anda mungkin juga menyukai
- Metabolisme Protein Dan LemakDokumen25 halamanMetabolisme Protein Dan LemakMulkan ArsyuddinBelum ada peringkat
- Energi BrutoDokumen5 halamanEnergi BrutoammiBelum ada peringkat
- Kebutuhan Zat Makanan Ayam Pedaging Dan PetelurDokumen29 halamanKebutuhan Zat Makanan Ayam Pedaging Dan PetelurLA CatHouseBelum ada peringkat
- Interelasi Metabolisme Dan Penentuan KaloriDokumen5 halamanInterelasi Metabolisme Dan Penentuan KaloriRomi Mauliza FauziBelum ada peringkat
- Bioenergitika 1Dokumen22 halamanBioenergitika 1alvian handiyantoBelum ada peringkat
- Resume Katabolisme Dan Anabolisme ProfesiDokumen87 halamanResume Katabolisme Dan Anabolisme ProfesiRisma SintyaBelum ada peringkat
- Energi Dalam Pakan RuminansiaDokumen26 halamanEnergi Dalam Pakan RuminansiaMarsetyo Bin SarjonoBelum ada peringkat
- Modul 2. EnergiDokumen18 halamanModul 2. EnergiDion TefaBelum ada peringkat
- EnergiDokumen35 halamanEnergiRetnoCahyaMuktiBelum ada peringkat
- Energi Untuk RuminansiaDokumen13 halamanEnergi Untuk RuminansiaUmmiBelum ada peringkat
- Bab V Nutrisi IkanDokumen81 halamanBab V Nutrisi Ikandinda putri amaliaBelum ada peringkat
- Pemanfaatan Energi MakananDokumen16 halamanPemanfaatan Energi MakananFrrlBhanu MBelum ada peringkat
- Energi Pada Ternak RuminansiaDokumen24 halamanEnergi Pada Ternak RuminansiaMarsetyo Bin Sarjono100% (1)
- MODUL 1 NUT224 Metabolisme Energi Dan Zat Gizi Makro Dudung AngkasaDokumen16 halamanMODUL 1 NUT224 Metabolisme Energi Dan Zat Gizi Makro Dudung AngkasaInstalasi Gizi RSUD PakuhajiBelum ada peringkat
- Metabolisme Pembentukan EnergiDokumen53 halamanMetabolisme Pembentukan EnergiNdy AgustaviaBelum ada peringkat
- ENERGIDokumen39 halamanENERGIDewi HandayaniBelum ada peringkat
- Pemanfaatan Energi Dalam Tubuh IkanDokumen3 halamanPemanfaatan Energi Dalam Tubuh IkanMulkan ArsyuddinBelum ada peringkat
- BioenergetikaDokumen5 halamanBioenergetikaAde MulkiBelum ada peringkat
- Keseimbangan EnergiDokumen19 halamanKeseimbangan EnergiYAn Na IsmiBelum ada peringkat
- Kandungan Energi MakananDokumen2 halamanKandungan Energi MakananrosaBelum ada peringkat
- Keseimbangan Energi Dan MetabolismeDokumen45 halamanKeseimbangan Energi Dan MetabolismeCynthiaSeptiantiBelum ada peringkat
- LP Dan Kasus Gangguan Pemenuhan Kebutuhan NutrisiDokumen22 halamanLP Dan Kasus Gangguan Pemenuhan Kebutuhan Nutrisiimran pasharBelum ada peringkat
- BioenergetikaDokumen27 halamanBioenergetika058-SULASTRI PRIHANDINIBelum ada peringkat
- Nutrisi Ikan - 4Dokumen35 halamanNutrisi Ikan - 4Dr.Ir. Subandiyono, MAppSc.75% (4)
- Bioenergetik Nutrisi Pada IkanDokumen20 halamanBioenergetik Nutrisi Pada Ikananna najjilil aniBelum ada peringkat
- Kebutuhan Energi-1Dokumen19 halamanKebutuhan Energi-1AyuseniBelum ada peringkat
- LAPORAN PENDAHULUAN NutrisiDokumen17 halamanLAPORAN PENDAHULUAN NutrisiShintia Andriani83% (18)
- Energi Dan NutrisiDokumen7 halamanEnergi Dan NutrisividyaBelum ada peringkat
- Bioenergetika Menggambarkan Aliran Energi Dan Nutrisi Dalam Sistem Biologi Dalam Contoh KitaDokumen13 halamanBioenergetika Menggambarkan Aliran Energi Dan Nutrisi Dalam Sistem Biologi Dalam Contoh KitaEzie Fp100% (1)
- Makalah DemamDokumen15 halamanMakalah DemamFrandy Huang100% (1)
- Pang4223 M1Dokumen58 halamanPang4223 M1Gea PalaBelum ada peringkat
- LP NutrisiDokumen17 halamanLP NutrisichrisnaBelum ada peringkat
- EnergiDokumen26 halamanEnergiAkbar UytBelum ada peringkat
- Istilah MetabolismeDokumen3 halamanIstilah MetabolismeYHulaiisoeun SaranghaeBeastelfshawol YoodoogiwoonjunseungBelum ada peringkat
- Ada DehDokumen8 halamanAda DehwilldanBelum ada peringkat
- Fungsi Makanan Bagi Tubuh ManusiaDokumen29 halamanFungsi Makanan Bagi Tubuh Manusianofile100% (1)
- Keseimbangan Energi Pengaturan Suhu TubuhDokumen6 halamanKeseimbangan Energi Pengaturan Suhu TubuhCharles Patrice Isaupu GansangBelum ada peringkat
- Metabolisme VITAMIN CDokumen25 halamanMetabolisme VITAMIN CMustapa Saputra PurbaBelum ada peringkat
- Uas Fisiologi OlahragaDokumen5 halamanUas Fisiologi OlahragaAju Al-GhifaryBelum ada peringkat
- Metabolisme Dan Keseimbangan EnergiDokumen5 halamanMetabolisme Dan Keseimbangan EnergiExma Mu'tatal HikmahBelum ada peringkat
- Metabolisme Zat Gizi (Resume)Dokumen13 halamanMetabolisme Zat Gizi (Resume)renyBelum ada peringkat
- Soal + Jawaban Ujian BioenergetikaDokumen12 halamanSoal + Jawaban Ujian BioenergetikaNining Cie PhalosaBelum ada peringkat
- Adaptasi Metabolik Pada LatihanDokumen14 halamanAdaptasi Metabolik Pada LatihanAminuddin NyampoBelum ada peringkat
- Modul Jadi Gizi OlahragaDokumen113 halamanModul Jadi Gizi OlahragaAgustin Dwi ErlandiBelum ada peringkat
- Asuhan Keperawatan Pada Pemenuhan Kebutuhan NutrisiDokumen25 halamanAsuhan Keperawatan Pada Pemenuhan Kebutuhan NutrisiSalsabilaBelum ada peringkat
- Metabolisme Dan Pengaturan Suhu TubuhDokumen17 halamanMetabolisme Dan Pengaturan Suhu TubuhVhero Tololiu100% (2)
- Apakah Itu Keseimbangan EnergiDokumen7 halamanApakah Itu Keseimbangan Energidhea1angela1putriBelum ada peringkat
- Laporan Pendahuluan4Dokumen8 halamanLaporan Pendahuluan4almaBelum ada peringkat
- LP NutrisiDokumen15 halamanLP NutrisiwisriBelum ada peringkat
- Energi Dan Laju MetabolismeDokumen17 halamanEnergi Dan Laju MetabolismeDiar AbdussalamBelum ada peringkat
- Makalah Sistem Pencernaan Pada HewanDokumen23 halamanMakalah Sistem Pencernaan Pada HewanMarkus Iyus SupiandiBelum ada peringkat
- Buku Masakan untuk Penyembuhan Skoliosis Anda: Jadikan tulang belakang lebih sehat dengan mengatur apa yang anda makan!Dari EverandBuku Masakan untuk Penyembuhan Skoliosis Anda: Jadikan tulang belakang lebih sehat dengan mengatur apa yang anda makan!Penilaian: 3.5 dari 5 bintang3.5/5 (4)
- Teknik Asas Pembiakan Hibrid Ikan Keli Thailand (Clarias sp.)Dari EverandTeknik Asas Pembiakan Hibrid Ikan Keli Thailand (Clarias sp.)Penilaian: 5 dari 5 bintang5/5 (2)
- Seksual SekunderDokumen5 halamanSeksual SekunderRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- Tugas IndividuDokumen7 halamanTugas IndividuRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- Tugas IndividuDokumen6 halamanTugas IndividuRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- Paper Praktek LapangDokumen8 halamanPaper Praktek LapangRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- Ikan BelanakDokumen5 halamanIkan BelanakRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- ArtemiaDokumen11 halamanArtemiaRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- I Mun Osti MulanDokumen11 halamanI Mun Osti MulanRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- Tugas Kelompok Udang GalahDokumen13 halamanTugas Kelompok Udang GalahRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- LAPORANDokumen18 halamanLAPORANRanhii'siiepoetriforget Maaharaniiy0% (1)
- My Xo BacteriaDokumen10 halamanMy Xo BacteriaRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- Makalah SelDokumen21 halamanMakalah SelRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- Fosfat TanahDokumen12 halamanFosfat TanahRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- Laporan Histologi Rani RevisiDokumen15 halamanLaporan Histologi Rani RevisiRanhii'siiepoetriforget MaaharaniiyBelum ada peringkat
- Makalah Pemupukan Dan PengapuranDokumen16 halamanMakalah Pemupukan Dan PengapuranRanhii'siiepoetriforget Maaharaniiy67% (3)
- Tekstur TanahDokumen15 halamanTekstur TanahRanhii'siiepoetriforget Maaharaniiy100% (1)