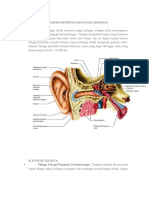
Anda mungkin juga menyukai
- Fisiologi Sistem PendengaranDokumen9 halamanFisiologi Sistem PendengaranArtrinda AnggitaBelum ada peringkat
- Anatomi TelingaDokumen35 halamanAnatomi TelingaHaqiqi Amira SyathirBelum ada peringkat
- Fisiologi PendengaranDokumen15 halamanFisiologi PendengaranAnnisa KartikasariBelum ada peringkat
- Makalah Kelompok Auditori Bab 1Dokumen13 halamanMakalah Kelompok Auditori Bab 1Zulkipli Maulana100% (1)
- Telinga Merupakan Sebuah Organ Yang Mampu MendeteksiDokumen23 halamanTelinga Merupakan Sebuah Organ Yang Mampu MendeteksiIsnaini AmaliahBelum ada peringkat
- Telinga Merupakan Salah Satu Indera Penting Bagi Kehidupan Manusia Veldi BaruDokumen6 halamanTelinga Merupakan Salah Satu Indera Penting Bagi Kehidupan Manusia Veldi BaruFera SalasiwaBelum ada peringkat
- StrukturDokumen12 halamanStrukturAmat Si DjibrilBelum ada peringkat
- Fisiologi Sistem PendengaranDokumen11 halamanFisiologi Sistem PendengaranJames Garcia50% (2)
- Laporan Kelompok Tutorial Ske 1 Blok THTDokumen45 halamanLaporan Kelompok Tutorial Ske 1 Blok THTSausan Hana MaharaniBelum ada peringkat
- Anatomi Telinga (Pendengaran) Ns NURHIDAYAHDokumen32 halamanAnatomi Telinga (Pendengaran) Ns NURHIDAYAHAldry UmagapBelum ada peringkat
- Skenario 2Dokumen22 halamanSkenario 2Koko KikoBelum ada peringkat
- Laporan Anatomi Histologi Fisiologi Ray Modul 2 SSDokumen10 halamanLaporan Anatomi Histologi Fisiologi Ray Modul 2 SSImam FahrizalBelum ada peringkat
- PBL Panca Indera Tuli PerseptifDokumen11 halamanPBL Panca Indera Tuli Perseptifniszamaf0% (1)
- Kul 2 &10 Sistem Pendengaran 2Dokumen112 halamanKul 2 &10 Sistem Pendengaran 2Erina NopiyantiBelum ada peringkat
- Referat AudiometriDokumen8 halamanReferat AudiometriAnastasia Caesarani KristelBelum ada peringkat
- Anfis PendengaranDokumen7 halamanAnfis PendengaranRias Sri UtamiBelum ada peringkat
- Proses PendengaranDokumen8 halamanProses PendengaranSuhaila Zakiya NajahBelum ada peringkat
- Laporan Fisiologi PendengaranDokumen5 halamanLaporan Fisiologi Pendengarandhia adhi perwirawatiBelum ada peringkat
- Auditory Processing DisorderDokumen24 halamanAuditory Processing Disorderindriyanti natasya ayu utami kottenBelum ada peringkat
- Praktikum Bioakustik GarputalaDokumen13 halamanPraktikum Bioakustik GarputalaNadia ChannelBelum ada peringkat
- Anatomi Telinga ManusiaDokumen14 halamanAnatomi Telinga ManusiaRiska WinarniBelum ada peringkat
- Fisiologi Sistem PendengaranDokumen26 halamanFisiologi Sistem PendengaranArizal SetyawanBelum ada peringkat
- Perhitungan Cacat PendengaranDokumen15 halamanPerhitungan Cacat PendengaranagungBelum ada peringkat
- TM SK2 A7 9057 Denis Dida FahrezziDokumen27 halamanTM SK2 A7 9057 Denis Dida FahrezziDenis DidaBelum ada peringkat
- Fisiologi PendengaranDokumen9 halamanFisiologi PendengaranFafelia RMBelum ada peringkat
- Biofisika (Pendengaran)Dokumen28 halamanBiofisika (Pendengaran)Finni Aulia SariBelum ada peringkat
- Modul 6 - Kegiatan Praktikum 3 - Percobaan Mekanisme Transmisi PendengaranDokumen4 halamanModul 6 - Kegiatan Praktikum 3 - Percobaan Mekanisme Transmisi PendengaranListari WasinoBelum ada peringkat
- Indra PendengaranDokumen19 halamanIndra Pendengarankartika SuciBelum ada peringkat
- Biokimia Indera PendengaranDokumen4 halamanBiokimia Indera PendengaranAnggina Mita Amalia0% (1)
- Anatomi TelingaDokumen31 halamanAnatomi Telingapandu putraBelum ada peringkat
- 7.1 PendengaranDokumen3 halaman7.1 Pendengaranarni rizha lentiniBelum ada peringkat
- Tuli SensorineuralDokumen9 halamanTuli SensorineuralSartika Putri31Belum ada peringkat
- Laporan Praktikum Faal PendengaranDokumen25 halamanLaporan Praktikum Faal PendengarantegarauliaBelum ada peringkat
- Bio Akustik 1Dokumen30 halamanBio Akustik 1ester zulfanisBelum ada peringkat
- Praktikum Kepekaan Indrea Pendengaran ManusiaDokumen10 halamanPraktikum Kepekaan Indrea Pendengaran ManusiaYulaekah ZulleBelum ada peringkat
- Anfis - Patofisiologi Penyakit - Indera TelingaDokumen59 halamanAnfis - Patofisiologi Penyakit - Indera Telingasiasat 400Belum ada peringkat
- Transmisi Gelombang SuaraDokumen2 halamanTransmisi Gelombang SuaraJordhi ValentinoBelum ada peringkat
- Laporan Ipa Kepekaan IndraDokumen9 halamanLaporan Ipa Kepekaan IndraJanahBelum ada peringkat
- Tugas 7 - Siti Rohmah - 2202171013 - Telinga Dan BagiannyaDokumen5 halamanTugas 7 - Siti Rohmah - 2202171013 - Telinga Dan BagiannyaSiti RohmahBelum ada peringkat
- Alps 2Dokumen4 halamanAlps 2firdaBelum ada peringkat
- Pemeriksaan Audiometri DasarDokumen43 halamanPemeriksaan Audiometri DasarRia Tutkey-MelmambessyBelum ada peringkat
- BioakustikDokumen32 halamanBioakustikftasitiBelum ada peringkat
- Pemeriksaan Tes PendengaranDokumen17 halamanPemeriksaan Tes PendengaranCindokeyBelum ada peringkat
- Laporan Fisiologi Blok XviiiDokumen16 halamanLaporan Fisiologi Blok XviiiHenry IrawanBelum ada peringkat
- Laporan Praktikum Biologi Perambatan Bunyi Melalui Tulang TengkorakDokumen22 halamanLaporan Praktikum Biologi Perambatan Bunyi Melalui Tulang TengkorakSofyan Dwi NugrohoBelum ada peringkat
- Perambatan Bunyi Melalui Tulang TengkorakDokumen14 halamanPerambatan Bunyi Melalui Tulang TengkorakIsmayanti NursiyamBelum ada peringkat
- NIHLDokumen8 halamanNIHLRizka_aprilBelum ada peringkat
- Indera Pendengar ManusiaDokumen3 halamanIndera Pendengar Manusiaupin ipinBelum ada peringkat
- KKPMT Kelompok 5 (Pendengaran)Dokumen10 halamanKKPMT Kelompok 5 (Pendengaran)Arif NugrohoBelum ada peringkat
- SerumenDokumen14 halamanSerumenGhinna Septhiana PratiwiBelum ada peringkat
- Fisika Telinga Dan PendengaranDokumen38 halamanFisika Telinga Dan PendengaranPuspa ArmanditaBelum ada peringkat
- Catatan Kuliah Anatomi TelingaDokumen24 halamanCatatan Kuliah Anatomi TelingaAdhityas Angga KusumaBelum ada peringkat
- Praktikum Alat PendengaranDokumen28 halamanPraktikum Alat PendengaranerizadeadaraBelum ada peringkat
- Indera Pendengaran Pada ManusiaDokumen9 halamanIndera Pendengaran Pada Manusiamelinda SilalahiBelum ada peringkat
- Anatomi Fisiologi TelingaDokumen7 halamanAnatomi Fisiologi TelingajuventuspengkulBelum ada peringkat
- Proses Produksi Suara ManusiaDokumen6 halamanProses Produksi Suara ManusiaEri NuraidaBelum ada peringkat
- Rangkuman Responsi TelingaDokumen25 halamanRangkuman Responsi TelingaPreston MitchellBelum ada peringkat
- Buku Kosakata Bahasa Jerman: Pendekatan Berbasis TopikDari EverandBuku Kosakata Bahasa Jerman: Pendekatan Berbasis TopikBelum ada peringkat
- Buku Kosakata Bahasa Suomi: Pendekatan Berbasis TopikDari EverandBuku Kosakata Bahasa Suomi: Pendekatan Berbasis TopikBelum ada peringkat
- Anti Remed Skenario 2Dokumen6 halamanAnti Remed Skenario 2Emír Gemilang JayaringgaBelum ada peringkat
- Modul Blok 8 Kongenital, Tumor Dan DegeneratifDokumen25 halamanModul Blok 8 Kongenital, Tumor Dan DegeneratifEmír Gemilang JayaringgaBelum ada peringkat
- Rat Sat Re Pemb Terpadu TKDokumen13 halamanRat Sat Re Pemb Terpadu TKchalkzzz100% (1)
- Skenario Blok 7 - SIstem Reproduksi Dan UrinariaDokumen3 halamanSkenario Blok 7 - SIstem Reproduksi Dan UrinariaEmír Gemilang JayaringgaBelum ada peringkat
- SISTEMEKSKRESIPADAMANUSIADokumen8 halamanSISTEMEKSKRESIPADAMANUSIADevi Siti ShofiyahBelum ada peringkat
- Pengumuman DamkarDokumen2 halamanPengumuman DamkarEmír Gemilang JayaringgaBelum ada peringkat
- Bola BasketDokumen6 halamanBola BasketEmír Gemilang JayaringgaBelum ada peringkat