Anda mungkin juga menyukai
- Pedoman Nasional Tatalaksana Klinis Infeksi HIV Dan Terapi Antiretroviral Pada Orang DewasaDokumen108 halamanPedoman Nasional Tatalaksana Klinis Infeksi HIV Dan Terapi Antiretroviral Pada Orang DewasaKeyla Kehara Putri100% (4)
- MiopiaDokumen20 halamanMiopiaInten Nur Rasadina100% (1)
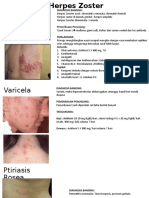
- Herpes Vs DermatitisDokumen38 halamanHerpes Vs DermatitisWika'deKaka'UtomoBelum ada peringkat
- Kumpulan Soal Latihan Bimo 3Dokumen117 halamanKumpulan Soal Latihan Bimo 3Siti HalimahBelum ada peringkat
- Kumpulan Soal Latihan Bimo 3Dokumen117 halamanKumpulan Soal Latihan Bimo 3Siti HalimahBelum ada peringkat
- Dermatologi Diagnosis dan Tatalaksana SingkatDokumen26 halamanDermatologi Diagnosis dan Tatalaksana SingkatTasya Sabrina Chairunnisa100% (1)
- Farmakokinetik Dan Farmakodinamik MeropenemDokumen7 halamanFarmakokinetik Dan Farmakodinamik MeropenemMaria FernandezBelum ada peringkat
- Anti MikrobaDokumen83 halamanAnti MikrobalanangBelum ada peringkat
- Kode Error Ir 5000-6000Dokumen16 halamanKode Error Ir 5000-6000toni jawarmanBelum ada peringkat
- Komputasi Rekayasa MaterialDokumen20 halamanKomputasi Rekayasa MaterialbabaBelum ada peringkat
- Laporan Praktikum Kimia Komputasi Modul 5-6Dokumen10 halamanLaporan Praktikum Kimia Komputasi Modul 5-6Dina nurkaniawatiBelum ada peringkat
- Laporan Praktikum Kimia Komputasi Modul 4Dokumen7 halamanLaporan Praktikum Kimia Komputasi Modul 4Dina nurkaniawatiBelum ada peringkat
- Modul 8: Hubungan Kuantitatif Struktur Aktivitas (HKSA) - 3Dokumen5 halamanModul 8: Hubungan Kuantitatif Struktur Aktivitas (HKSA) - 3Thoriq NurdinBelum ada peringkat
- Digital Canon Error CodesDokumen3 halamanDigital Canon Error CodesTriwija KesumaBelum ada peringkat
- Kerusakan TVDokumen27 halamanKerusakan TVhaerBelum ada peringkat
- Canon Ir 5000 inDokumen12 halamanCanon Ir 5000 inNindya Saksana100% (1)
- LAPORAN PRAKTIKUMDokumen14 halamanLAPORAN PRAKTIKUMMa YLBelum ada peringkat
- Laporan Praktikum Kimia Komputasi Modul 3Dokumen7 halamanLaporan Praktikum Kimia Komputasi Modul 3Dina nurkaniawatiBelum ada peringkat
- Kode Error Fotocopy Canon IR 6570 Bahasa Indonesia - Mesin FotoDokumen4 halamanKode Error Fotocopy Canon IR 6570 Bahasa Indonesia - Mesin FotoRamsi Ankzi100% (1)
- Kinetika Reaksi Ion Permanganat Dengan ADokumen19 halamanKinetika Reaksi Ion Permanganat Dengan APradipta NovitasariBelum ada peringkat
- tUGAS Kmo (Comfa Comsia)Dokumen3 halamantUGAS Kmo (Comfa Comsia)rosyti100% (1)
- Kode Galat Mesin Foto CopyDokumen14 halamanKode Galat Mesin Foto CopyExco EndarBelum ada peringkat
- Kode Error Ir5075Dokumen1 halamanKode Error Ir5075Fauzan AzymaBelum ada peringkat
- TranskripsiDokumen35 halamanTranskripsiGema Masse CiptaBelum ada peringkat
- FORMULIR PENDAFTARAN MHS STMNUKWAR 2023Dokumen1 halamanFORMULIR PENDAFTARAN MHS STMNUKWAR 2023SEKOLAH TINGGI SAINS DAN TEKNOLOGI MANOKWARIBelum ada peringkat
- Cara Menggunakan Network ScanGearDokumen16 halamanCara Menggunakan Network ScanGearVideo GoodBelum ada peringkat
- Makalah BiosensorDokumen12 halamanMakalah BiosensoriishariyantiBelum ada peringkat
- Regulasi EnzimDokumen11 halamanRegulasi EnzimArdjani 09Belum ada peringkat
- Belajar Linux MudahDokumen6 halamanBelajar Linux MudahtuffrohuBelum ada peringkat
- Bahan Diskusi Bio 2Dokumen3 halamanBahan Diskusi Bio 2Nastiti Apsari AjiduniBelum ada peringkat
- Gelombang dan PartikelDokumen18 halamanGelombang dan PartikelWulantikaSintiaBelum ada peringkat
- Error Code Ir3300 Ir2800 Ir2200Dokumen7 halamanError Code Ir3300 Ir2800 Ir2200Mudi YonoBelum ada peringkat
- Catkul Prof SadikinDokumen10 halamanCatkul Prof SadikinSisca Lioe0% (1)
- Buku Kimia Organik SintesisDokumen58 halamanBuku Kimia Organik Sintesisshofiatul aliyaBelum ada peringkat
- MDRDokumen10 halamanMDRbangkit99Belum ada peringkat
- Resistensi Obat MalariaDokumen22 halamanResistensi Obat MalariaChandraAstiBelum ada peringkat
- Dr. Triwani - Deteksi Resistensi Plasmodium Falciparum Terhadap Klorokuin Dengan Marka Situs Polimorfik Lys76Tyr Gen PFCRT Menggunakan PCR-RFDokumen8 halamanDr. Triwani - Deteksi Resistensi Plasmodium Falciparum Terhadap Klorokuin Dengan Marka Situs Polimorfik Lys76Tyr Gen PFCRT Menggunakan PCR-RFBudi LamakaBelum ada peringkat
- MEKANISME RESISTENSI MALARIADokumen13 halamanMEKANISME RESISTENSI MALARIASari0% (1)
- Terapi Malaria Pada KehamilanDokumen11 halamanTerapi Malaria Pada Kehamilannurulnadya100% (2)
- Resistensi Pengobatan Anti MalariaDokumen14 halamanResistensi Pengobatan Anti MalariaRosiana AryaniBelum ada peringkat
- Faktor Genetik yang Mempengaruhi Respon Obat WarfarinDokumen6 halamanFaktor Genetik yang Mempengaruhi Respon Obat WarfarinPuspita WsrBelum ada peringkat
- Tugas BM SelvianaDokumen2 halamanTugas BM SelvianaSelviana FebrianiBelum ada peringkat
- RIFAMPISIN REVIEWDokumen6 halamanRIFAMPISIN REVIEWLaila Vie VieLaBelum ada peringkat
- Obat RifampisinDokumen5 halamanObat RifampisinccnadiaBelum ada peringkat
- OBAT ANTIPROTOZOADokumen38 halamanOBAT ANTIPROTOZOAHamidahBelum ada peringkat
- Obat Malaria Mekanisme KerjaDokumen10 halamanObat Malaria Mekanisme KerjaqyuraBelum ada peringkat
- Obat Malaria Dan Mekanisme KerjanyaDokumen3 halamanObat Malaria Dan Mekanisme KerjanyaAlfriyHani MBelum ada peringkat
- Tugas Farmakoterapi IIDokumen7 halamanTugas Farmakoterapi IIFhY-thRee ZhafhYthree JazManBelum ada peringkat
- 2552 3254 1 PBDokumen7 halaman2552 3254 1 PBSri komalaBelum ada peringkat
- Polimorfisme Gen Cyp2c8 Dan Pengaruhnya Terhadap Terapi MalariaDokumen2 halamanPolimorfisme Gen Cyp2c8 Dan Pengaruhnya Terhadap Terapi MalariaKhotimatulBelum ada peringkat
- Goodman GilmanDokumen10 halamanGoodman GilmanGabriella StellyBelum ada peringkat
- Antivirus FarmakologiDokumen20 halamanAntivirus FarmakologiDesy AprianiBelum ada peringkat
- Pengaruh SLCO1B1 Terhadap RifampisinDokumen3 halamanPengaruh SLCO1B1 Terhadap RifampisinC4613 36 Warda Fatin NabilaBelum ada peringkat
- CYP2C8 MalariaDokumen5 halamanCYP2C8 MalariaAndang Wisnu SeptianBelum ada peringkat
- GEN POLIMORFISME MEMPENGARUHI RESPON OBATDokumen3 halamanGEN POLIMORFISME MEMPENGARUHI RESPON OBATI Kadek Adi Putra Suandana 2005Belum ada peringkat
- Keracunan KloramfenikolDokumen7 halamanKeracunan KloramfenikolHaudina Rahma KladiaBelum ada peringkat
- 15-Article Text-51-1-10-20180607Dokumen7 halaman15-Article Text-51-1-10-20180607Nedita Putri BandaroBelum ada peringkat
- MDR TB IndonisiaDokumen19 halamanMDR TB IndonisiaAnggri SeptyanBelum ada peringkat
- ANTIPROTOZOADokumen38 halamanANTIPROTOZOAKeerthy Venthen100% (1)
- IdDokumen8 halamanIdRIKABelum ada peringkat
- Tugas BiomolekulerDokumen7 halamanTugas BiomolekulerSparkling PinkBelum ada peringkat
- Jawaban Uts Lab Dasar DR - IrsanDokumen10 halamanJawaban Uts Lab Dasar DR - IrsansariBelum ada peringkat
- Peran Enzim Di Bidang FarmasiDokumen9 halamanPeran Enzim Di Bidang FarmasiRohani Panjaitan0% (1)
- KEKEBALAN NYAMUK TERHADAP PLASMODIUMDokumen11 halamanKEKEBALAN NYAMUK TERHADAP PLASMODIUMhanafi Bin SyafrizalBelum ada peringkat
- Polimorfisme Gen VKORC1 dan Pengaruhnya terhadap Aktivitas WarfarinDokumen13 halamanPolimorfisme Gen VKORC1 dan Pengaruhnya terhadap Aktivitas WarfarinI Kadek Adi Putra Suandana 2005Belum ada peringkat
- Tugas FarmakologiDokumen27 halamanTugas FarmakologiNesyaBelum ada peringkat
- Kuliah THTDokumen58 halamanKuliah THTTuan Muda TunggulBelum ada peringkat
- Kaidah Dasar BioetikaDokumen29 halamanKaidah Dasar BioetikaSiti HalimahBelum ada peringkat
- Respi 1Dokumen3 halamanRespi 1Siti HalimahBelum ada peringkat
- Bells PalsyDokumen8 halamanBells PalsyDewa SuryaBelum ada peringkat
- PPT VertigoDokumen33 halamanPPT VertigoSiti HalimahBelum ada peringkat
- Pretes Aipki2 BaruDokumen42 halamanPretes Aipki2 BaruSiti HalimahBelum ada peringkat
- Soal Pretest Batch NovDokumen49 halamanSoal Pretest Batch NovSiti HalimahBelum ada peringkat
- FolikulitisDokumen13 halamanFolikulitisreza auliaBelum ada peringkat
- Bells PalsyDokumen8 halamanBells PalsyDewa SuryaBelum ada peringkat
- Ruptum PerineumDokumen9 halamanRuptum PerineumSiti HalimahBelum ada peringkat
- E-Soal UKMPPD FORENSIK - OutDokumen20 halamanE-Soal UKMPPD FORENSIK - OutAfifah Nur RahmiBelum ada peringkat
- KP 19.21 Visus Refraksi - 2008Dokumen46 halamanKP 19.21 Visus Refraksi - 2008Ahmad ShukriBelum ada peringkat
- Syok AnafilaksisDokumen13 halamanSyok AnafilaksisMutiarawati MithrataelúwånzhùBelum ada peringkat
- Kusta - LepraDokumen25 halamanKusta - LepraSiti HalimahBelum ada peringkat
- OBAT - AipkiDokumen3 halamanOBAT - AipkiSiti HalimahBelum ada peringkat
- DETEKSI DINI TBDokumen17 halamanDETEKSI DINI TBfairuz roji akbarBelum ada peringkat
- DEFINISI ABORTUSDokumen56 halamanDEFINISI ABORTUSSiti HalimahBelum ada peringkat
- Malaria BaruDokumen16 halamanMalaria BaruSiti HalimahBelum ada peringkat
- Syok AnafilaksisDokumen13 halamanSyok AnafilaksisMutiarawati MithrataelúwånzhùBelum ada peringkat
- 8 48 1 PBDokumen8 halaman8 48 1 PBMayalisna PrihatiningrumBelum ada peringkat
- ISKDokumen11 halamanISKSiti HalimahBelum ada peringkat
- HIPERTENSI PRIMERDokumen14 halamanHIPERTENSI PRIMERSiti HalimahBelum ada peringkat
- HIPERTENSI PRIMERDokumen14 halamanHIPERTENSI PRIMERSiti HalimahBelum ada peringkat
- Empyema LengkapDokumen41 halamanEmpyema LengkapganispyBelum ada peringkat