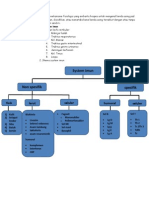
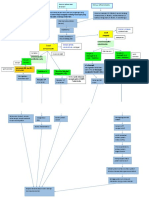
Anda mungkin juga menyukai
- Bab 2Dokumen4 halamanBab 2Trisna TRBelum ada peringkat
- Komplemen Sistem ImunDokumen13 halamanKomplemen Sistem ImunYulia AnggreniBelum ada peringkat
- Biologi MolekulerDokumen15 halamanBiologi MolekulerDiah Yulia CitraBelum ada peringkat
- PPT Bakteri Tahan AsamDokumen62 halamanPPT Bakteri Tahan Asamfirdausia roseBelum ada peringkat
- Naftyan YF (PCR Konvesional & Real Time)Dokumen8 halamanNaftyan YF (PCR Konvesional & Real Time)Eva MaulaBelum ada peringkat
- CRP 2Dokumen7 halamanCRP 2Medina Fadlilatus SyaadahBelum ada peringkat
- Kit Insert TranslateDokumen2 halamanKit Insert TranslateYopi NurhikmahBelum ada peringkat
- 2a - Kelompok 1 - Parasitologi - Coleoptera & HymenopteraDokumen18 halaman2a - Kelompok 1 - Parasitologi - Coleoptera & HymenopteraSeptiyanti EkaBelum ada peringkat
- Modifikasi Pemeriksaan LeukositDokumen45 halamanModifikasi Pemeriksaan LeukositKarolina Hutagalung100% (1)
- IMUNOSEROLOGIDokumen11 halamanIMUNOSEROLOGIMelina Rahman100% (1)
- SoalDokumen8 halamanSoalMatthew MckenzieBelum ada peringkat
- Deteksi Asam NukleatDokumen16 halamanDeteksi Asam NukleatAdlimatulPutriIlmiyahBelum ada peringkat
- Pemeriksaan HBsAgDokumen5 halamanPemeriksaan HBsAgMeikeBelum ada peringkat
- Sitti Mantasia-Ekstraksi & PCRDokumen60 halamanSitti Mantasia-Ekstraksi & PCRRahmawati RusmanBelum ada peringkat
- Tugas Imunoserologi P Musthari (Evi)Dokumen13 halamanTugas Imunoserologi P Musthari (Evi)drgirsangBelum ada peringkat
- Mekanisme Kerja Western BlotDokumen5 halamanMekanisme Kerja Western BlotPoe TraBelum ada peringkat
- Penanda Garis Keturunan Forensik Dan Aplikasi PCRDokumen16 halamanPenanda Garis Keturunan Forensik Dan Aplikasi PCRyuniBelum ada peringkat
- Kimia KlinikDokumen19 halamanKimia KlinikSyaharani RaniBelum ada peringkat
- Makalah Autoimunitas Atika Jaya Rani (13330716)Dokumen46 halamanMakalah Autoimunitas Atika Jaya Rani (13330716)SwerysBelum ada peringkat
- Uji Skrinning Teknik ImmunoassayDokumen17 halamanUji Skrinning Teknik ImmunoassayPUDJI HARDJANTOBelum ada peringkat
- Retraksi BekuanDokumen14 halamanRetraksi BekuanRinaRastutiBelum ada peringkat
- ElisaDokumen46 halamanElisapungky pradanaBelum ada peringkat
- Draft Modul Mikrobiologi - RevDokumen126 halamanDraft Modul Mikrobiologi - RevIgma TrisaBelum ada peringkat
- Laporan Praktek Lahan Imunoserologi IDokumen53 halamanLaporan Praktek Lahan Imunoserologi IHarto waldiBelum ada peringkat
- Boyolali 12 Jan 20. Kendali Mutu Dan Validasi ImunoserologiDokumen25 halamanBoyolali 12 Jan 20. Kendali Mutu Dan Validasi ImunoserologiendyBelum ada peringkat
- Sistem ImunDokumen4 halamanSistem ImunX-KaizenBelum ada peringkat
- Di Susun OlehDokumen18 halamanDi Susun OlehNirwanaBelum ada peringkat
- Laporan Praktikum Bakteriologi Iik BWDokumen17 halamanLaporan Praktikum Bakteriologi Iik BWDamascena CocleBelum ada peringkat
- Sop Cystatin C FixDokumen5 halamanSop Cystatin C FixRenanda AlifianBelum ada peringkat
- Quality Control - LabDokumen29 halamanQuality Control - Labdiahnisa7Belum ada peringkat
- 6.02 - Makalah Biologi Molekuler - Kelompok 9 - PCRDokumen12 halaman6.02 - Makalah Biologi Molekuler - Kelompok 9 - PCRSyafira ArifaBelum ada peringkat
- Toksikokinetik 2Dokumen6 halamanToksikokinetik 2yopaBelum ada peringkat
- File Soal Ukom Alcian BlueDokumen16 halamanFile Soal Ukom Alcian BluesaniaBelum ada peringkat
- Warna Tabung PemeriksaanDokumen6 halamanWarna Tabung PemeriksaanAdinda AzhafirahBelum ada peringkat
- CD 4 Adalah Sebuah Marker Atau Penanda Yang Berada Di Permukaan SelDokumen7 halamanCD 4 Adalah Sebuah Marker Atau Penanda Yang Berada Di Permukaan SelAllen ParmanBelum ada peringkat
- Kualitas Dna Hasil Isolasi Dari Beberapa Bagian BatangDokumen17 halamanKualitas Dna Hasil Isolasi Dari Beberapa Bagian BatangSiti KhoirunnisaBelum ada peringkat
- Uji Fungsi Hati SGPT SgotDokumen12 halamanUji Fungsi Hati SGPT SgotdyaBelum ada peringkat
- Pembahasan InfekDokumen4 halamanPembahasan InfekHendro Dwi SugiyantoBelum ada peringkat
- Latihan Soal FarmakologiDokumen6 halamanLatihan Soal FarmakologiVivian Yulias Sabela100% (1)
- Pemeriksaan HCGDokumen19 halamanPemeriksaan HCGDyahra TyasBelum ada peringkat
- Hitung Jenis LeukositDokumen4 halamanHitung Jenis LeukositSabila RosyidaBelum ada peringkat
- Diagnosa Laboratorium ShigellaDokumen3 halamanDiagnosa Laboratorium ShigellaDaniel WilliamsBelum ada peringkat
- Mikologi Kelompok 4Dokumen47 halamanMikologi Kelompok 4Salma NFBelum ada peringkat
- Laporan Praktikum SitohistoteknologiDokumen4 halamanLaporan Praktikum Sitohistoteknologiastrii 08Belum ada peringkat
- Petunjuk Praktikum PK - GITDokumen29 halamanPetunjuk Praktikum PK - GITkhairunnisaBelum ada peringkat
- 1188-Article Text-5613-1-10-20210828Dokumen66 halaman1188-Article Text-5613-1-10-20210828rossi akalBelum ada peringkat
- P01 001B Touchdown PCRDokumen3 halamanP01 001B Touchdown PCRsaadah fauziyahBelum ada peringkat
- Makalah Imunohematologi Dan Bank Darah PDFDokumen12 halamanMakalah Imunohematologi Dan Bank Darah PDFFitriza AfkaBelum ada peringkat
- Soal Bakteri Pat Xii 22 - 23 CBTDokumen7 halamanSoal Bakteri Pat Xii 22 - 23 CBTAgung SetiyawanBelum ada peringkat
- Toleransi ImunogenikDokumen11 halamanToleransi ImunogenikIsniNobilisMogaaUtamiBelum ada peringkat
- Hasil Pengamatan Hema SanglahDokumen13 halamanHasil Pengamatan Hema SanglahadiBelum ada peringkat
- Kimia KlinikDokumen14 halamanKimia KlinikEsda PotTer100% (1)
- 2020 2 Pengenalan Laboratorium PADokumen28 halaman2020 2 Pengenalan Laboratorium PAr. raisy100% (1)
- Laporan Praktikum Mikrobiologi 4F - Hasil Pengamatan Dan PembahasanDokumen2 halamanLaporan Praktikum Mikrobiologi 4F - Hasil Pengamatan Dan PembahasanAfdilah IrawatiBelum ada peringkat
- QC Imunoserologi PX Widal N CRPDokumen11 halamanQC Imunoserologi PX Widal N CRPWulan NurevaBelum ada peringkat
- Proto-Onkogen Dan OnkogenDokumen5 halamanProto-Onkogen Dan OnkogenSekar NugraheniBelum ada peringkat
- MakroamilasemiaDokumen1 halamanMakroamilasemiasyahdah iksiroh al husnahBelum ada peringkat
- IMLTD Donor DarahDokumen2 halamanIMLTD Donor Darahpenastinisa67% (3)
- Referat AutoimunDokumen56 halamanReferat AutoimunMaya Jou-san Ismayanti100% (1)
- Toleransi ImunDokumen20 halamanToleransi ImunSamuel FpBelum ada peringkat
- Skema Mekanisme Penyembuhan Mukosa OralDokumen3 halamanSkema Mekanisme Penyembuhan Mukosa OralsuhanaBelum ada peringkat
- Ob 5 Makalah Kelompok 1Dokumen4 halamanOb 5 Makalah Kelompok 1suhanaBelum ada peringkat
- Skema Patogenesis Penyakit Kelenjar SalivaDokumen3 halamanSkema Patogenesis Penyakit Kelenjar SalivasuhanaBelum ada peringkat
- Skema Patogenesis Penyakit Kelenjar SalivaDokumen3 halamanSkema Patogenesis Penyakit Kelenjar SalivasuhanaBelum ada peringkat
- Ob 5 Makalah Kelompok 1Dokumen4 halamanOb 5 Makalah Kelompok 1suhanaBelum ada peringkat
- Kurnia SelaDokumen25 halamanKurnia SelasuhanaBelum ada peringkat
- TMJ (Anatomi)Dokumen9 halamanTMJ (Anatomi)suhanaBelum ada peringkat