Anda mungkin juga menyukai
- Metabolisme Purin Dan PirimidinDokumen19 halamanMetabolisme Purin Dan PirimidinDanny Adi KurniawanBelum ada peringkat
- Metabolisme Asam Amino Dan ProteinDokumen22 halamanMetabolisme Asam Amino Dan ProteinRahmat Darma Wansyah100% (7)
- Zona Kamar BedahDokumen13 halamanZona Kamar BedahFahmiCullen100% (2)
- Asam NukleatDokumen15 halamanAsam NukleatSiskaJanerBelum ada peringkat
- Makalah METABOLISME NUKLEOTIDADokumen19 halamanMakalah METABOLISME NUKLEOTIDABaldus Pierre Bouvier67% (3)
- Metabolisme Asam Amino 2Dokumen29 halamanMetabolisme Asam Amino 2Adhia DekaBelum ada peringkat
- REPLIKASI, Transkripsi, Translasi Dna RnaDokumen11 halamanREPLIKASI, Transkripsi, Translasi Dna RnasenoBelum ada peringkat
- Makalah Biokimia - Metabolisme Asam NukleatDokumen14 halamanMakalah Biokimia - Metabolisme Asam NukleatMas RipanBelum ada peringkat
- Biosintesis NukleotidaDokumen7 halamanBiosintesis NukleotidarezaBelum ada peringkat
- Degradasi PurinDokumen8 halamanDegradasi PurinLukia Nabila100% (3)
- Makalah (Biosintesis Purin)Dokumen9 halamanMakalah (Biosintesis Purin)Adrianii Nha-Nya Haniie Nomleni0% (1)
- 4-Reaksi Kimia Asam AminoDokumen13 halaman4-Reaksi Kimia Asam AminoJaya PonselBelum ada peringkat
- Kelainan Metabolisme ProteinDokumen18 halamanKelainan Metabolisme ProteinTiak TuyonoBelum ada peringkat
- Makalah Biokimia Asam Nukleat PDFDokumen21 halamanMakalah Biokimia Asam Nukleat PDFFuad JanyBelum ada peringkat
- Biosintesis Asam AminoDokumen6 halamanBiosintesis Asam AminoMagnalia Morena RuthBelum ada peringkat
- Anabolisme LipidDokumen22 halamanAnabolisme LipidrossyBelum ada peringkat
- Latar Belakang Asam AminoDokumen8 halamanLatar Belakang Asam AminoGunawan Bastian NugratamaBelum ada peringkat
- Anabolisme LemakDokumen27 halamanAnabolisme LemakUnggul Teguh PrasetyoBelum ada peringkat
- Regulasi EnzimDokumen9 halamanRegulasi EnzimBilal SabikhismaBelum ada peringkat
- Struktur Asam NukleatDokumen14 halamanStruktur Asam NukleatJolteon JuniorBelum ada peringkat
- Biosintesis Asam LemakDokumen8 halamanBiosintesis Asam LemakanbBelum ada peringkat
- Makalah Asam NukleatDokumen18 halamanMakalah Asam NukleatFira WatiBelum ada peringkat
- Anabolisme LemakDokumen20 halamanAnabolisme LemakGita Dinia Putri67% (3)
- Asam LemakDokumen25 halamanAsam Lemakmeghamirrah100% (2)
- Pembentukan Ikatan PeptidaDokumen8 halamanPembentukan Ikatan Peptidataul75% (4)
- Katabolisme Asam AminoDokumen26 halamanKatabolisme Asam Aminolia mulyani100% (4)
- Klasifikasi KarbohidratDokumen6 halamanKlasifikasi Karbohidratinneke putri0% (1)
- Biokimia Biosintesis ProteinDokumen15 halamanBiokimia Biosintesis ProteinRisniBelum ada peringkat
- Dogma SentralDokumen6 halamanDogma SentralEko Budi NBelum ada peringkat
- Enzim Pada Metabolisme Lemak Di Sistem Pencernaan Dan Mekanisme KerjanyaDokumen13 halamanEnzim Pada Metabolisme Lemak Di Sistem Pencernaan Dan Mekanisme Kerjanyaumi_armansyahBelum ada peringkat
- Uji ProteinDokumen18 halamanUji ProteinEka WidiastutikBelum ada peringkat
- Alanin Saskhia LeonyDokumen9 halamanAlanin Saskhia LeonySaskhia GomezBelum ada peringkat
- Biokim Makalah-Biosintesis ProteinDokumen41 halamanBiokim Makalah-Biosintesis Proteinsyaadahfs100% (3)
- Asam Nukleat, Asnawar - A22120140 Kls EDokumen9 halamanAsam Nukleat, Asnawar - A22120140 Kls EAsnawar RustamBelum ada peringkat
- p3 - Reaksi Uji Protein KitaDokumen23 halamanp3 - Reaksi Uji Protein KitassuuzzaannBelum ada peringkat
- Transaminasi DeaminasiDokumen3 halamanTransaminasi DeaminasiFransiska Anggitha71% (7)
- BIOKIMIA Nukleotida NukleosidaDokumen65 halamanBIOKIMIA Nukleotida NukleosidaSriSumartini0% (1)
- Pilihan Ganda Soal Jawaban Genetika Dan Sintesis Protein 20 ButirDokumen7 halamanPilihan Ganda Soal Jawaban Genetika Dan Sintesis Protein 20 ButirIntan SoerayaBelum ada peringkat
- Tinjauan Pustaka RNADokumen4 halamanTinjauan Pustaka RNAIkhwanMaulanaBelum ada peringkat
- Presentasi Anabolisme LipidDokumen41 halamanPresentasi Anabolisme LipidTerrina ChairulitaBelum ada peringkat
- Anabolisme LipidDokumen55 halamanAnabolisme LipidLala Febrianti100% (4)
- Mekanisme Perbaikan DnaDokumen4 halamanMekanisme Perbaikan Dnajamardi sinagaBelum ada peringkat
- Metabolisme LipidDokumen33 halamanMetabolisme LipidYuniar33% (3)
- Glikogenesis Adalah Proses Pembentukan Atau Biosintesis Glikogen Yang Terjadi Terutama Di Dalam Hati Dan OtotDokumen2 halamanGlikogenesis Adalah Proses Pembentukan Atau Biosintesis Glikogen Yang Terjadi Terutama Di Dalam Hati Dan OtotWindi SetianyBelum ada peringkat
- Metabolisme Nukleotida Purin Dan Pirimidin RepairedDokumen21 halamanMetabolisme Nukleotida Purin Dan Pirimidin RepairedFathimatuz Zahro0% (1)
- Biokimia TRANSLASIDokumen18 halamanBiokimia TRANSLASIAiny Kusuma100% (1)
- Makalah Protein Asam AminoDokumen19 halamanMakalah Protein Asam AminoAnggun LestariBelum ada peringkat
- Penentuan Kadar ProteinDokumen29 halamanPenentuan Kadar ProteinKhudrotunnada AdityaBelum ada peringkat
- Uji Identifikasi ProteinDokumen16 halamanUji Identifikasi ProteinAwhine Chen0% (2)
- Pengertian Dan Definisi Asam NukleatDokumen32 halamanPengertian Dan Definisi Asam NukleatNur Hidayat33% (3)
- Biosintesis LipidDokumen6 halamanBiosintesis Lipidakhmad11350% (2)
- Fosforilasi OksidatifDokumen21 halamanFosforilasi OksidatifAkhi Manan SetiawanBelum ada peringkat
- Uji Asam AminoDokumen19 halamanUji Asam AminoLuciana Mentari HutapeaBelum ada peringkat
- Makalah TranslasiDokumen10 halamanMakalah Translasishadow_anchorBelum ada peringkat
- Pembahasan Uji Koagulasi & DenaturasiDokumen9 halamanPembahasan Uji Koagulasi & DenaturasiIndarti Ulfayani0% (1)
- Kelompok 1 Asam NukleatDokumen15 halamanKelompok 1 Asam NukleatFaizul MaknunBelum ada peringkat
- MAKALAH BIOTEKNOLOGI FARMASI Asam Nukleat Dan Protein Alih InformasiDokumen24 halamanMAKALAH BIOTEKNOLOGI FARMASI Asam Nukleat Dan Protein Alih InformasidonicrusoeBelum ada peringkat
- Resume Asam Nukleat Regina Cyahyani P. PaputunganDokumen11 halamanResume Asam Nukleat Regina Cyahyani P. PaputunganRegina cyahyani PaputunganBelum ada peringkat
- Nukleotida Dan Asam NukleatDokumen20 halamanNukleotida Dan Asam NukleatIlham Surya AbadiBelum ada peringkat
- Makalah Biokimia Asam NukleatDokumen34 halamanMakalah Biokimia Asam NukleatNurrillahBelum ada peringkat
- Makalah Biokimia Umum II - Metabolisme Protein Dan Asam NukleatDokumen29 halamanMakalah Biokimia Umum II - Metabolisme Protein Dan Asam NukleatMas RipanBelum ada peringkat
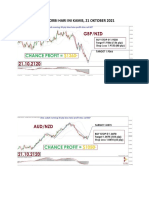
- Analisa 21 Oktober 2021Dokumen2 halamanAnalisa 21 Oktober 2021Christianto PasunuBelum ada peringkat
- Undang-Undang No 30 Tahun 2002 Tentang Komisi Pemberantasan KorupsiDokumen6 halamanUndang-Undang No 30 Tahun 2002 Tentang Komisi Pemberantasan KorupsiChristianto PasunuBelum ada peringkat
- R P P Operasi Bentuk AljabarDokumen7 halamanR P P Operasi Bentuk AljabarDea Putri TariBelum ada peringkat
- Buku Anti KorupsiDokumen59 halamanBuku Anti KorupsiChristianto PasunuBelum ada peringkat
- Bab 3Dokumen4 halamanBab 3Christianto PasunuBelum ada peringkat
- Analisa 19 Oktober 2021Dokumen7 halamanAnalisa 19 Oktober 2021Christianto PasunuBelum ada peringkat
- Makalah Bayi Tabung FinishDokumen20 halamanMakalah Bayi Tabung FinishAnisa Maulina50% (2)
- Tumbuh Kembang Anak 3Dokumen17 halamanTumbuh Kembang Anak 3Christianto PasunuBelum ada peringkat
- Tumbuh Kembang Anak Melly2Dokumen6 halamanTumbuh Kembang Anak Melly2Christianto PasunuBelum ada peringkat
- Zener HotspotDokumen1 halamanZener HotspotChristianto PasunuBelum ada peringkat
- Full Skripsi NisaDokumen54 halamanFull Skripsi NisaChristianto PasunuBelum ada peringkat
- R P P Operasi Bentuk AljabarDokumen7 halamanR P P Operasi Bentuk AljabarDea Putri TariBelum ada peringkat
- Tumbuh Kembang Anak Melly2Dokumen6 halamanTumbuh Kembang Anak Melly2Christianto PasunuBelum ada peringkat
- Abs TrakDokumen1 halamanAbs TrakChristianto PasunuBelum ada peringkat
- Abs TrakDokumen1 halamanAbs TrakChristianto PasunuBelum ada peringkat
- Hubungan Tingkat Kecemasan Dengan MekanismeDokumen16 halamanHubungan Tingkat Kecemasan Dengan MekanismeChristianto PasunuBelum ada peringkat
- BAGIANDDokumen89 halamanBAGIANDChristianto PasunuBelum ada peringkat
- Untuk Pembina PSDokumen2 halamanUntuk Pembina PSChristianto PasunuBelum ada peringkat
- Luas LingkaranDokumen6 halamanLuas LingkaranChristianto PasunuBelum ada peringkat
- Bayi Tabung Menurut Pandangan KristianiDokumen13 halamanBayi Tabung Menurut Pandangan KristianiYeni100% (2)
- Bahan PenelitianDokumen1 halamanBahan PenelitianChristianto PasunuBelum ada peringkat
- Data LogbookDokumen1 halamanData LogbookChristianto PasunuBelum ada peringkat
- Penge Sah AnDokumen1 halamanPenge Sah AnChristianto PasunuBelum ada peringkat
- Tugas Stabilitas ObatDokumen6 halamanTugas Stabilitas ObatChristianto PasunuBelum ada peringkat
- Sampul OrsinDokumen1 halamanSampul OrsinChristianto PasunuBelum ada peringkat
- Kerangka Acuan LEADINGDokumen6 halamanKerangka Acuan LEADINGChristianto PasunuBelum ada peringkat
- On Mipa 2014Dokumen15 halamanOn Mipa 2014Himapemia Fkip UnjaBelum ada peringkat
- Jurnal EmulsiDokumen6 halamanJurnal EmulsiChristianto Pasunu100% (1)