Anda mungkin juga menyukai
- Jurnal Belajar HemichordataDokumen12 halamanJurnal Belajar HemichordataLelly Luckitasari100% (1)
- Asterias OphiothrixDokumen16 halamanAsterias OphiothrixayBelum ada peringkat
- Biologi PerairanDokumen13 halamanBiologi PerairanFarras FaishalBelum ada peringkat
- Anatomi SefalokordataDokumen5 halamanAnatomi Sefalokordataindriana megaBelum ada peringkat
- Makalah Cephalochordata 1-1Dokumen10 halamanMakalah Cephalochordata 1-1Winda KanukBelum ada peringkat
- Lampyran - Texture Batuan BekuDokumen21 halamanLampyran - Texture Batuan BekuVilman SidikBelum ada peringkat
- Kelas AgnathaDokumen30 halamanKelas AgnathaArman Abu Bakar100% (2)
- Rahma Novianti - 1304620033 - Laporan Praktikum Filum AnnelidaDokumen10 halamanRahma Novianti - 1304620033 - Laporan Praktikum Filum AnnelidaRahma NoviantiBelum ada peringkat
- Filum ChordataDokumen20 halamanFilum ChordataDesyAAlvridaBelum ada peringkat
- PolychaetaDokumen56 halamanPolychaetaM Adnan Kurnianto100% (1)
- Echinodermata OoDokumen49 halamanEchinodermata OomufidaBelum ada peringkat
- Hemi CordataDokumen50 halamanHemi CordataTiara Dwi NurmalitaBelum ada peringkat
- MoluscaDokumen17 halamanMoluscaadrianaBelum ada peringkat
- AnnelidaDokumen28 halamanAnnelidanurul lailiBelum ada peringkat
- Laporan Lengkap ReptilDokumen10 halamanLaporan Lengkap ReptilAndi Sry Fifin AndrianiBelum ada peringkat
- Latihan IVDokumen19 halamanLatihan IVMuhammad Ma'arij JayaBelum ada peringkat
- Bab 8 Kingdom AnimaliaDokumen92 halamanBab 8 Kingdom AnimaliaSalsabilla Kimiko100% (1)
- Makalah CephalochordataDokumen19 halamanMakalah Cephalochordatasheilla100% (1)
- EchinodermataDokumen12 halamanEchinodermataikmal maulasanyBelum ada peringkat
- ChordataDokumen30 halamanChordataMiftahurrohmat Almaasyah100% (1)
- Zoovert KLP 10Dokumen5 halamanZoovert KLP 10nurBelum ada peringkat
- Bahan CestodaDokumen42 halamanBahan CestodaPebrian Diki PrestyaBelum ada peringkat
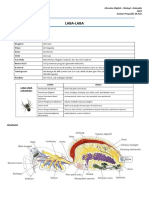
- Laba LabaDokumen13 halamanLaba LabawarungmocoBelum ada peringkat
- Fisiologi Filum MolluscaDokumen9 halamanFisiologi Filum MolluscaadekBelum ada peringkat
- Zoologi 1Dokumen5 halamanZoologi 1BilqisBelum ada peringkat
- Asteroidea OphiuroideaDokumen7 halamanAsteroidea Ophiuroideaida nurpitaBelum ada peringkat
- MolluscaDokumen22 halamanMolluscanadianandariniBelum ada peringkat
- Animalia (Dunia Hewan)Dokumen48 halamanAnimalia (Dunia Hewan)Finaya Ailsa SetyaBelum ada peringkat
- Blok 7 Up 5Dokumen14 halamanBlok 7 Up 5Meyla CitraBelum ada peringkat
- Bab 6. Ashelminthes: 1.1. Filum GastrotrichaDokumen14 halamanBab 6. Ashelminthes: 1.1. Filum GastrotrichaDita Azzahra Fara DisaBelum ada peringkat
- Bab 9 AnnelidaDokumen41 halamanBab 9 AnnelidaAan TilolangoBelum ada peringkat
- Laporan Praktikum Mamalia Kelompok 5. RevDokumen13 halamanLaporan Praktikum Mamalia Kelompok 5. RevKadek Wirna Dewi SuaningsihBelum ada peringkat
- Cakupan Materi UrochordataDokumen6 halamanCakupan Materi UrochordataAstri anggrianiBelum ada peringkat
- CEFALOAGNATADokumen42 halamanCEFALOAGNATADinarAjengNurAzizaBelum ada peringkat
- Ordo OrthopteraDokumen15 halamanOrdo OrthopterariashaniaBelum ada peringkat
- ReptiliaDokumen38 halamanReptiliaRismaAfridaRosaniaBelum ada peringkat
- Acara 11 - Organ Dan Sistem Organ Pada AmphibiaDokumen13 halamanAcara 11 - Organ Dan Sistem Organ Pada AmphibiaEster Feberiana100% (1)
- Mollusca Kelompok ArinaDokumen25 halamanMollusca Kelompok ArinaARNYBelum ada peringkat
- Dasar Teori MolluscaDokumen21 halamanDasar Teori MolluscaNugraha Saputra SBelum ada peringkat
- PiscesDokumen24 halamanPiscesDarwin azis100% (1)
- Sub Filum UrochordataDokumen6 halamanSub Filum Urochordataisfatun chasanah100% (2)
- ArthropodaDokumen9 halamanArthropodafajriBelum ada peringkat
- MoluscaDokumen7 halamanMoluscaMewa MahartaBelum ada peringkat
- Sistem Respirasi InvertebrataDokumen21 halamanSistem Respirasi InvertebratariniaBelum ada peringkat
- Bahan Ajar InvertebrataDokumen153 halamanBahan Ajar InvertebrataNovita RahayuBelum ada peringkat
- Jurnal Belajar 3Dokumen15 halamanJurnal Belajar 3Ardiani Samti Nur AzizahBelum ada peringkat
- Ordo Siphonaptera - DR - DinaDokumen14 halamanOrdo Siphonaptera - DR - DinaRamaSumualBelum ada peringkat
- Bab Ii PrintDokumen22 halamanBab Ii PrintwandiBelum ada peringkat
- Laporan Praktikum Mollusca (Biologi)Dokumen18 halamanLaporan Praktikum Mollusca (Biologi)Anggii WidyBelum ada peringkat
- Resume AnnelidaDokumen10 halamanResume Annelidanaimmatus0% (1)
- Makalah Chondrichtyes 2-1Dokumen21 halamanMakalah Chondrichtyes 2-1ilmisaBelum ada peringkat
- Makalah OsteichthyesDokumen21 halamanMakalah Osteichthyesjoniwijayaaa88% (8)
- DokumenDokumen5 halamanDokumenYudha WidiastikaBelum ada peringkat
- Klasifikasi Ikan Kakap MerahDokumen17 halamanKlasifikasi Ikan Kakap MerahichsanBelum ada peringkat
- Makalah CrustaceaDokumen24 halamanMakalah CrustaceaAhmad Happy M S100% (5)
- PRAKTIKUM VI MolluscaDokumen27 halamanPRAKTIKUM VI MolluscaMuhammad Luthfi33% (3)
- 09 AnnelidaDokumen41 halaman09 Annelidapuskesmas moutongBelum ada peringkat
- Famili AcanthamoebidaeDokumen22 halamanFamili Acanthamoebidaean964Belum ada peringkat
- Kelas Conoidasida, Subclass GregarinasinaDokumen5 halamanKelas Conoidasida, Subclass Gregarinasinaan964Belum ada peringkat
- Amino Tingoensefalitis Primer (PAM) .Dokumen8 halamanAmino Tingoensefalitis Primer (PAM) .an964Belum ada peringkat
- Trematoda DigeneansDokumen9 halamanTrematoda Digeneansan964Belum ada peringkat
- Trematoda AspidobothreaDokumen12 halamanTrematoda Aspidobothreaan964Belum ada peringkat
- Identifikasi-Larva & PupaDokumen16 halamanIdentifikasi-Larva & Pupaan964Belum ada peringkat