Anda mungkin juga menyukai
- Mikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaDari EverandMikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaPenilaian: 2.5 dari 5 bintang2.5/5 (2)
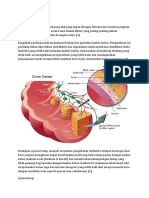
- Tinjauan Pustaka Ca ColonDokumen19 halamanTinjauan Pustaka Ca Colonnirmala amirBelum ada peringkat
- Mikrobiologi Medis I: Patogen dan Mikrobioma ManusiaDari EverandMikrobiologi Medis I: Patogen dan Mikrobioma ManusiaPenilaian: 4 dari 5 bintang4/5 (11)
- Epidemiologi KankerDokumen19 halamanEpidemiologi KankerApri NingsihBelum ada peringkat
- Coronavirus Covid-19. Membela diri. Cara menghindari penularan. Bagaimana melindungi keluarga dan pekerjaan Anda. Diperbarui edisi keempat.Dari EverandCoronavirus Covid-19. Membela diri. Cara menghindari penularan. Bagaimana melindungi keluarga dan pekerjaan Anda. Diperbarui edisi keempat.Penilaian: 5 dari 5 bintang5/5 (2)
- Penatalaksanaan PTG Resiko TinggiDokumen7 halamanPenatalaksanaan PTG Resiko TinggiYusrizal Adnan100% (1)
- Teknik Asas Pembiakan Hibrid Ikan Keli Thailand (Clarias sp.)Dari EverandTeknik Asas Pembiakan Hibrid Ikan Keli Thailand (Clarias sp.)Penilaian: 5 dari 5 bintang5/5 (2)
- Contoh TugasDokumen27 halamanContoh TugasuliyantiBelum ada peringkat
- Panduan Esensial untuk Skoliosis dan Kesehatan KehamilanDari EverandPanduan Esensial untuk Skoliosis dan Kesehatan KehamilanPenilaian: 4.5 dari 5 bintang4.5/5 (4)
- Kanker Pada Usia LanjutDokumen18 halamanKanker Pada Usia LanjutJennie FpBelum ada peringkat
- Buku Masakan untuk Penyembuhan Skoliosis Anda: Jadikan tulang belakang lebih sehat dengan mengatur apa yang anda makan!Dari EverandBuku Masakan untuk Penyembuhan Skoliosis Anda: Jadikan tulang belakang lebih sehat dengan mengatur apa yang anda makan!Penilaian: 3.5 dari 5 bintang3.5/5 (4)
- Askep CA Ovarium (Stgo)Dokumen18 halamanAskep CA Ovarium (Stgo)Elke Catur100% (1)
- Laporan Pendahuluan CA RectiDokumen40 halamanLaporan Pendahuluan CA RectiCintya AdiantiBelum ada peringkat
- Sindrom MetabolikDokumen38 halamanSindrom MetabolikEvan Marpaung100% (1)
- Penyakit-Trofoblas-Gestasional 45Dokumen28 halamanPenyakit-Trofoblas-Gestasional 45mochamad rizaBelum ada peringkat
- Laporan Sitostatik CA ColorektalDokumen26 halamanLaporan Sitostatik CA ColorektalHerlina AlfianyBelum ada peringkat
- Penyakit Trofoblas GestasionalDokumen25 halamanPenyakit Trofoblas GestasionalDennys KerehBelum ada peringkat
- KMB - Kelompok 2B. Analisa Laporan CA Gaster-1Dokumen36 halamanKMB - Kelompok 2B. Analisa Laporan CA Gaster-1Rizkycia Chahya MorgaBelum ada peringkat
- Ca ColonDokumen71 halamanCa ColonChristina Airyunda PertiwiBelum ada peringkat
- Referat - Tumor ColonDokumen37 halamanReferat - Tumor Colonmuhamad isafarudinBelum ada peringkat
- Laporan PendahuluanDokumen12 halamanLaporan PendahuluanDianita SafitriBelum ada peringkat
- LP CA CaecumDokumen16 halamanLP CA Caecum'Mbem Octaviani'saiank Ndud-foreverBelum ada peringkat
- Ca MammaeDokumen16 halamanCa Mammaeklinik unilaBelum ada peringkat
- Bab II CA KolonDokumen14 halamanBab II CA KolonRukmanaBelum ada peringkat
- Referat Penyakit Trofoblas GanasDokumen29 halamanReferat Penyakit Trofoblas GanasSherry KingBelum ada peringkat
- Kanker KolorektalDokumen8 halamanKanker KolorektalSeto Wahyu PrasetyoBelum ada peringkat
- Kanker Kolorektal AlomedikaDokumen16 halamanKanker Kolorektal AlomedikaWilky WaluyoBelum ada peringkat
- Ca MammaeDokumen5 halamanCa MammaealdianaBelum ada peringkat
- Ca ColonDokumen42 halamanCa ColonNurul SimatupangBelum ada peringkat
- Makalah Kanker KolonDokumen23 halamanMakalah Kanker KolonMuh Ramadhan SaputroBelum ada peringkat
- Penatalaksanaan PTG Resiko TinggiDokumen7 halamanPenatalaksanaan PTG Resiko TinggifitriaBelum ada peringkat
- ASUHAN KEPERAWATAN PADA PASIEN DENGAN CA OVARIUM Dock RARADokumen11 halamanASUHAN KEPERAWATAN PADA PASIEN DENGAN CA OVARIUM Dock RARArahmat dmiBelum ada peringkat
- Makalah Kanker Kolon by Siska AngrainiDokumen24 halamanMakalah Kanker Kolon by Siska AngrainiLeni PuspitasariBelum ada peringkat
- Laporan Pendahuluan CA. RECTIDokumen32 halamanLaporan Pendahuluan CA. RECTIDila Amalina ArselanBelum ada peringkat
- Asuhan Gizi Kanker Payudara - Kel 12BDokumen21 halamanAsuhan Gizi Kanker Payudara - Kel 12BFathya KhadijahBelum ada peringkat
- Referat Tumor Intra Abdomen-AlvinaDokumen28 halamanReferat Tumor Intra Abdomen-AlvinaAlvina DjoedirBelum ada peringkat
- Laporan Pendahuluan CA RECTIDokumen28 halamanLaporan Pendahuluan CA RECTIAvc SubangBelum ada peringkat
- LAPORAN PENDAHULUAN Ca RektiDokumen26 halamanLAPORAN PENDAHULUAN Ca Rektiyulione vickyBelum ada peringkat
- LP Tumor Intra AbdomenDokumen21 halamanLP Tumor Intra AbdomenRiyanti RiaBelum ada peringkat
- Askep Paliatif CA Ovarium RevisiDokumen31 halamanAskep Paliatif CA Ovarium RevisiMenny NovianiBelum ada peringkat
- KIA - Novy Maulid DianaDokumen56 halamanKIA - Novy Maulid DianaArsenio RaffaBelum ada peringkat
- Kanker ColonDokumen15 halamanKanker ColonMutia Farah DianaBelum ada peringkat
- LP Tumor Int AbdDokumen29 halamanLP Tumor Int Abdfidyaangraeni 61Belum ada peringkat
- LP Tu Intra AbdomenDokumen20 halamanLP Tu Intra Abdomeniqlima nafiisahBelum ada peringkat
- Laporan Pendahuluan Tumor Intra AbdomenDokumen18 halamanLaporan Pendahuluan Tumor Intra Abdomenrhadiatul67% (3)
- LP Ca RectiDokumen28 halamanLP Ca RectiAniezah Ku100% (2)
- LP - Tumor ColonDokumen21 halamanLP - Tumor ColonAgustin Tamara100% (1)
- Laporan Tutorial Skenerio 5 GUDokumen62 halamanLaporan Tutorial Skenerio 5 GUd_destungBelum ada peringkat
- LP Ca Recti. Anis WahyuDokumen19 halamanLP Ca Recti. Anis WahyuAnnizahro Nurrul SafitriBelum ada peringkat
- LP CA RectalDokumen26 halamanLP CA RectalRaka Fachrizal HilmyBelum ada peringkat
- LP Ca Colon MiraDokumen10 halamanLP Ca Colon MiraVennyta SariBelum ada peringkat
- Epidemiologi KankerDokumen68 halamanEpidemiologi Kankerm rizki akbarBelum ada peringkat
- Laporan Pendahuluan Klien Dengan CA RectiDokumen26 halamanLaporan Pendahuluan Klien Dengan CA RectiMary Peterson67% (3)
- Bio Assignmnet CancerDokumen8 halamanBio Assignmnet Cancernor ainBelum ada peringkat
- Makalah Kelompok 3 (CA Colon)Dokumen13 halamanMakalah Kelompok 3 (CA Colon)Vira AdeliaBelum ada peringkat
- LP Ca Recti - LP 2Dokumen28 halamanLP Ca Recti - LP 2qqh6bfvrtkBelum ada peringkat
- Ulasan: Anaplastic Thyroid Carcinoma: Diagnosis Dan Pengobatan Saat IniDokumen11 halamanUlasan: Anaplastic Thyroid Carcinoma: Diagnosis Dan Pengobatan Saat IniAulia AgmaBelum ada peringkat
- Sap CA ColonDokumen9 halamanSap CA ColonMaimunah RahmawatiBelum ada peringkat
- Laporan LBM 3 (SGD 8)Dokumen29 halamanLaporan LBM 3 (SGD 8)NitaBelum ada peringkat
- Kanker OvariumDokumen17 halamanKanker OvariumEmir Fakhrudin100% (1)
- SGD KonstipasiDokumen6 halamanSGD KonstipasiRusmalinda Septiana HasanahBelum ada peringkat
- Patofisiologi Ulkus DekubitusDokumen2 halamanPatofisiologi Ulkus DekubitusRusmalinda Septiana HasanahBelum ada peringkat
- SGD Geriatri KonstipasiDokumen2 halamanSGD Geriatri KonstipasiRusmalinda Septiana HasanahBelum ada peringkat
- Tatalaksana Konstipasi Pada LansiaDokumen2 halamanTatalaksana Konstipasi Pada LansiaRusmalinda Septiana HasanahBelum ada peringkat
- Patogenesis Dan Faktor Resiko KonstipasiDokumen3 halamanPatogenesis Dan Faktor Resiko KonstipasiRusmalinda Septiana HasanahBelum ada peringkat
- VaksinDokumen3 halamanVaksinRusmalinda Septiana HasanahBelum ada peringkat
- VaksinDokumen3 halamanVaksinRusmalinda Septiana HasanahBelum ada peringkat
- VaksinDokumen3 halamanVaksinRusmalinda Septiana HasanahBelum ada peringkat
- VaksinDokumen3 halamanVaksinRusmalinda Septiana HasanahBelum ada peringkat
- VaksinDokumen3 halamanVaksinRusmalinda Septiana HasanahBelum ada peringkat
- VaksinDokumen3 halamanVaksinRusmalinda Septiana HasanahBelum ada peringkat
- Boarding Pass Anatomi 4Dokumen3 halamanBoarding Pass Anatomi 4Rusmalinda Septiana HasanahBelum ada peringkat
- VaksinDokumen3 halamanVaksinRusmalinda Septiana HasanahBelum ada peringkat
- Patofisiologi StrokeDokumen2 halamanPatofisiologi StrokeRusmalinda Septiana HasanahBelum ada peringkat
- Tatalaksana Tinea UnguiumDokumen6 halamanTatalaksana Tinea UnguiumRusmalinda Septiana HasanahBelum ada peringkat
- TL RWN-TL Dws-Proses-Sendi'15-6Dokumen76 halamanTL RWN-TL Dws-Proses-Sendi'15-6Rusmalinda Septiana HasanahBelum ada peringkat
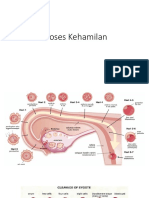
- Proses Kehamilan Mulai Dari PembuahanDokumen5 halamanProses Kehamilan Mulai Dari PembuahanRusmalinda Septiana HasanahBelum ada peringkat
- BALOKDokumen22 halamanBALOKRusmalinda Septiana HasanahBelum ada peringkat
- Gambaran Klinis AbortusDokumen4 halamanGambaran Klinis AbortusRusmalinda Septiana HasanahBelum ada peringkat
- MuscleDokumen27 halamanMuscleRusmalinda Septiana HasanahBelum ada peringkat