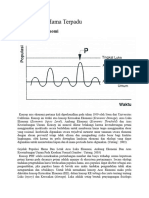
Anda mungkin juga menyukai
- Mikrobiologi Medis I: Patogen dan Mikrobioma ManusiaDari EverandMikrobiologi Medis I: Patogen dan Mikrobioma ManusiaPenilaian: 4 dari 5 bintang4/5 (11)
- Mikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaDari EverandMikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaPenilaian: 2.5 dari 5 bintang2.5/5 (2)
- Pengendalian Hama Secara HayatiDokumen20 halamanPengendalian Hama Secara HayatiIlham Inzide Corp100% (1)
- Pengendalian Hama Terpadu (Isi)Dokumen13 halamanPengendalian Hama Terpadu (Isi)Anisa RahmalitaBelum ada peringkat
- BAB IV Budidaya Bawang MerahDokumen9 halamanBAB IV Budidaya Bawang MerahmilaBelum ada peringkat
- Konsep Aras EkonomiDokumen19 halamanKonsep Aras EkonomiEta Fanani AR100% (1)
- Laporan Besar DPT (L2)Dokumen27 halamanLaporan Besar DPT (L2)Dwi Yan NovitBelum ada peringkat
- Praktikum Acara 3 Penentuan Ambang EkonomiDokumen8 halamanPraktikum Acara 3 Penentuan Ambang Ekonomisulistiyanto0% (1)
- Makalah TPHDokumen13 halamanMakalah TPHmilaBelum ada peringkat
- Pengendalian Penyakit TugasDokumen13 halamanPengendalian Penyakit TugasNurhayati YusufBelum ada peringkat
- Konsep Ambang EkonomiDokumen19 halamanKonsep Ambang EkonomiAmos KorneliusBelum ada peringkat
- Dokumen 4-DikonversiDokumen4 halamanDokumen 4-DikonversiFanstagramBelum ada peringkat
- Konsep Aras Luka Ekonomi EİLDokumen7 halamanKonsep Aras Luka Ekonomi EİLAgustin ZarkaniBelum ada peringkat
- Riska Aristi - 2014131041 - Tugas DDPT BioekonomiDokumen2 halamanRiska Aristi - 2014131041 - Tugas DDPT BioekonomiRiska AristiBelum ada peringkat
- Ambang Ekonomi Pengendalian HamaDokumen23 halamanAmbang Ekonomi Pengendalian Hamaadrian100% (1)
- Pengelolaan Hama TerpaduDokumen3 halamanPengelolaan Hama TerpaduYafi Sahdha100% (1)
- Pemanfaatan Ambang Ekonomi Dalam Pelaksanaan PHT (Tamara Pantow 19031109019)Dokumen9 halamanPemanfaatan Ambang Ekonomi Dalam Pelaksanaan PHT (Tamara Pantow 19031109019)Alan Briwanly PelleBelum ada peringkat
- Dasar Pertimbangan Pengendalian Penyakit TanamanDokumen22 halamanDasar Pertimbangan Pengendalian Penyakit TanamanNdhRegBelum ada peringkat
- Laporan IHT BawangDokumen9 halamanLaporan IHT BawangRieyo SoesiloBelum ada peringkat
- Pengendalian Hama Dan Penyakit Secara TerpaduDokumen22 halamanPengendalian Hama Dan Penyakit Secara TerpaduEka Yuli SusantiBelum ada peringkat
- Aplikasi Herbisida 22Dokumen20 halamanAplikasi Herbisida 22Netanya PanggabeanBelum ada peringkat
- Dasar Perlindungan Tanaman RevisiDokumen13 halamanDasar Perlindungan Tanaman RevisiagastyaarbiBelum ada peringkat
- Tingkat Kerusakan Ekonomi Atau Ambang KerusakanDokumen11 halamanTingkat Kerusakan Ekonomi Atau Ambang Kerusakanyuyun lheaBelum ada peringkat
- Revisi Lap MHPTDokumen25 halamanRevisi Lap MHPTAhmad MundirBelum ada peringkat
- Segitiga PenyakitDokumen24 halamanSegitiga PenyakitRifki A. Fuady100% (1)
- Review Jurnal 6Dokumen5 halamanReview Jurnal 6DESTY CHINTHA HUKIBelum ada peringkat
- Laporan DPTDokumen56 halamanLaporan DPTAnnisa SetiyaraniBelum ada peringkat
- Ambang Ekonomi, Insidensi, Dan InsensitasDokumen5 halamanAmbang Ekonomi, Insidensi, Dan InsensitasHanifah IzzatiBelum ada peringkat
- Aras Keputusan Ekonomi Pengendalian Hama (7) 8Dokumen27 halamanAras Keputusan Ekonomi Pengendalian Hama (7) 8masmanira100% (1)
- Dasar Perlindungan TanamanDokumen6 halamanDasar Perlindungan TanamanMahrani 123Belum ada peringkat
- Metode Pengamatan Opt Tanaman BuahDokumen46 halamanMetode Pengamatan Opt Tanaman BuahFathurrahman Siddiq33% (3)
- Bottom-Up Dan Top-Down Effect Pada TAnaman Inang Terhadap Pola Interaksi Serangga Herbivora-Predatornya Pada Ekosistem Sawah-Penelitian Fundamental2013 - 2Dokumen3 halamanBottom-Up Dan Top-Down Effect Pada TAnaman Inang Terhadap Pola Interaksi Serangga Herbivora-Predatornya Pada Ekosistem Sawah-Penelitian Fundamental2013 - 2Ita FitriyanaBelum ada peringkat
- PHPT Kelompok 4Dokumen2 halamanPHPT Kelompok 4M Ilham AbrarBelum ada peringkat
- Ilmu Hama & PenyakitDokumen121 halamanIlmu Hama & PenyakitHendrik Al-FarisiBelum ada peringkat
- Tugas Hama Terpadu Andi Syahdila Herliana 1940201038Dokumen5 halamanTugas Hama Terpadu Andi Syahdila Herliana 1940201038Lia Kemba100% (1)
- Tugas Ambang Ekonomi IconDokumen3 halamanTugas Ambang Ekonomi IconCelineBelum ada peringkat
- 10 - Resume Pengendalian Hama - Tbio2Dokumen29 halaman10 - Resume Pengendalian Hama - Tbio2Putrii PuspiitaBelum ada peringkat
- Proposal AsmadiDokumen27 halamanProposal AsmadiDika SaputraBelum ada peringkat
- Hama Utama Kacang TanahDokumen20 halamanHama Utama Kacang Tanahmufida chairunisaBelum ada peringkat
- Pengelolaan Agroekosistem Dalam Pengendalian Opt Berdasarkan Konsep Pengendalian HayatiDokumen14 halamanPengelolaan Agroekosistem Dalam Pengendalian Opt Berdasarkan Konsep Pengendalian HayatiElvrado Wega SBelum ada peringkat
- DPT LamraDokumen15 halamanDPT LamraYOSUA YOUNGKIBelum ada peringkat
- Uts DPTDokumen3 halamanUts DPTIlham SolahuddinBelum ada peringkat
- Laporan PHPT LengkapDokumen32 halamanLaporan PHPT Lengkapfitri primaBelum ada peringkat
- Pengendalian Hama TerpaduDokumen20 halamanPengendalian Hama TerpaduAdi TamaBelum ada peringkat
- Dasar Perlindungan Tanaman Tugas KelompokDokumen7 halamanDasar Perlindungan Tanaman Tugas KelompokNADYA WIDI PRAMESWARIBelum ada peringkat
- Hubungan Populasi Dengan Tingkat SeranganDokumen6 halamanHubungan Populasi Dengan Tingkat SeranganArum B2utyBelum ada peringkat
- Pengendalian Hayati GulmaDokumen54 halamanPengendalian Hayati GulmaTuning Ridha AddhinyBelum ada peringkat
- Kurniyawan - Tugas DDPTDokumen2 halamanKurniyawan - Tugas DDPTKurniyawan KurniyawanBelum ada peringkat
- Ambang Batas Ekonomi Dalam Pengendalian Hama TerpaduDokumen14 halamanAmbang Batas Ekonomi Dalam Pengendalian Hama TerpaduIshom HadiqBelum ada peringkat
- Risiko Usaha Tani Bawang MerahDokumen5 halamanRisiko Usaha Tani Bawang MerahLink FaBelum ada peringkat
- Makalah DDA Kelompok 3Dokumen15 halamanMakalah DDA Kelompok 3Miya AdektriBelum ada peringkat
- ProteksiTanaman Alpukat Dari Serangan Jamur Tular Tanah Dengan Menggunakan Biofungisida Trichoderma HarzianumDokumen20 halamanProteksiTanaman Alpukat Dari Serangan Jamur Tular Tanah Dengan Menggunakan Biofungisida Trichoderma Harzianumropiq azisBelum ada peringkat
- Management) Merupakan Langkah Yang Sangat Strategis Dalam Kerangka TuntutanDokumen11 halamanManagement) Merupakan Langkah Yang Sangat Strategis Dalam Kerangka Tuntutansri ningsihBelum ada peringkat
- Ambang EkonomiDokumen2 halamanAmbang EkonomiNovelina SidaurukBelum ada peringkat
- 23-Article Text-91-1-10-20200829Dokumen7 halaman23-Article Text-91-1-10-20200829dunia sophieBelum ada peringkat
- Pengukuran Penyakit Dan Kehilangan HasilDokumen26 halamanPengukuran Penyakit Dan Kehilangan Hasilhartati oktarinaBelum ada peringkat
- Penggunaan Jamur Dan Bakteri PDFDokumen6 halamanPenggunaan Jamur Dan Bakteri PDFNaufalYaafiMurtadhoBelum ada peringkat
- Surat Perintah KerjaDokumen2 halamanSurat Perintah KerjamilaBelum ada peringkat
- Tumpangsari Jagung Dan Kacang TanahDokumen11 halamanTumpangsari Jagung Dan Kacang TanahmilaBelum ada peringkat
- TPH Makalah Pengendalian HayatiDokumen32 halamanTPH Makalah Pengendalian HayatimilaBelum ada peringkat
- Pengertian Pengendalian HayatiDokumen19 halamanPengertian Pengendalian HayatimilaBelum ada peringkat
- Acara IiiDokumen5 halamanAcara IiimilaBelum ada peringkat
- ACARA II VegetatifDokumen9 halamanACARA II VegetatifmilaBelum ada peringkat
- Lily PutihDokumen6 halamanLily PutihmilaBelum ada peringkat
- Acara ViDokumen14 halamanAcara VimilaBelum ada peringkat
- Bab I Identifikasi Dan Pemilihan Alatmesin Usaha Tani-1Dokumen13 halamanBab I Identifikasi Dan Pemilihan Alatmesin Usaha Tani-1milaBelum ada peringkat
- Bab I Identifikasi Dan Pemilihan Alatmesin Usaha Tani-1Dokumen13 halamanBab I Identifikasi Dan Pemilihan Alatmesin Usaha Tani-1milaBelum ada peringkat
- BAB III Budidaya Kubis Merah2Dokumen8 halamanBAB III Budidaya Kubis Merah2milaBelum ada peringkat
- Bab II Pengenalan Traktor Sebagai Sumber Tenaga, Cara Menyiapkan Dan Cara Mengemudikannya-1Dokumen18 halamanBab II Pengenalan Traktor Sebagai Sumber Tenaga, Cara Menyiapkan Dan Cara Mengemudikannya-1mila67% (3)
- Makalah Tekben Benih RekalsitranDokumen12 halamanMakalah Tekben Benih RekalsitranmilaBelum ada peringkat
- 1 Pengend Hayati PDFDokumen17 halaman1 Pengend Hayati PDFmilaBelum ada peringkat
- Acara IvDokumen17 halamanAcara Ivmila0% (1)