Anda mungkin juga menyukai
- Makalah Anatomi Dan Fisologi Perkembangan Mahkul HidupDokumen12 halamanMakalah Anatomi Dan Fisologi Perkembangan Mahkul Hidupnovia damaiBelum ada peringkat
- Laprak TatanamaDokumen26 halamanLaprak Tatanamamelania citraBelum ada peringkat
- Makalah Cryptogamae Kel IiDokumen23 halamanMakalah Cryptogamae Kel Iijuliana heldyBelum ada peringkat
- Tata NamaDokumen13 halamanTata NamaIrma YenniBelum ada peringkat
- Makalah BioumDokumen33 halamanMakalah Bioumshynta devita reginaBelum ada peringkat
- Taksonomi TranslateDokumen22 halamanTaksonomi TranslateNadaBelum ada peringkat
- Klasifikasi OrganismeDokumen39 halamanKlasifikasi OrganismeEru Triyanda NapitupuluBelum ada peringkat
- Studi Dalam Nomenklatur Dan KlasifikasiDokumen5 halamanStudi Dalam Nomenklatur Dan KlasifikasiAinur RahmatBelum ada peringkat
- Petunjuk Praktikum Taksonomi Tumbuhan 2015Dokumen39 halamanPetunjuk Praktikum Taksonomi Tumbuhan 2015Muladi JufriBelum ada peringkat
- Translate JurnalDokumen9 halamanTranslate JurnalMaydiana PutriBelum ada peringkat
- Jurnal Angiospermae 3.en - IdDokumen20 halamanJurnal Angiospermae 3.en - Idnanda anisaBelum ada peringkat
- Kesalahan Teori EvolusiDokumen16 halamanKesalahan Teori Evolusi100_wahyuniBelum ada peringkat
- TaxonomyDokumen37 halamanTaxonomyTeni Setia MulyaniBelum ada peringkat
- Tatanama HewanDokumen28 halamanTatanama Hewanrika ayunta100% (1)
- Makalah - Kelompok 1 - Kelas E - Prinsip Dan Tata Nama HewanDokumen11 halamanMakalah - Kelompok 1 - Kelas E - Prinsip Dan Tata Nama HewanDikaikaBelum ada peringkat
- Petunjuk Praktikum Taksonomi Tumbuhan 2016Dokumen39 halamanPetunjuk Praktikum Taksonomi Tumbuhan 2016AnuBelum ada peringkat
- Perbedaan Antara Taksonomi Dan KlasifikasiDokumen2 halamanPerbedaan Antara Taksonomi Dan KlasifikasiNurlaili Dwi Putri UmacinaBelum ada peringkat
- ProkariotikDokumen23 halamanProkariotikNadaBelum ada peringkat
- Materi 7 BioinformatikaDokumen11 halamanMateri 7 BioinformatikaAndini DwiyantiBelum ada peringkat
- Diversitas Dan Klasifikasi MikrobaDokumen10 halamanDiversitas Dan Klasifikasi Mikrobariska raharjoBelum ada peringkat
- 1 Pendahuluan Sistematika Tumbuhan IIDokumen23 halaman1 Pendahuluan Sistematika Tumbuhan IIasettodexterBelum ada peringkat
- M 2Dokumen12 halamanM 2Ade Kurnia RachmadaniBelum ada peringkat
- Nomenklatur KEL 6Dokumen16 halamanNomenklatur KEL 6Mutia SariBelum ada peringkat
- Modul Praktikum Sistematika TumbuhanDokumen34 halamanModul Praktikum Sistematika TumbuhanNirmala F. HabibieBelum ada peringkat
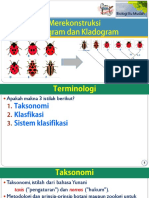
- Rekonstruksi Fenogram - Kladogram - Fotosintesis - 3Dokumen71 halamanRekonstruksi Fenogram - Kladogram - Fotosintesis - 3Rendi AdityaBelum ada peringkat
- ICBNDokumen23 halamanICBNHendrawan OyzBelum ada peringkat
- Makalah Ipa TerpaduDokumen29 halamanMakalah Ipa Terpadugebiselan54Belum ada peringkat
- Kode Internasional NomenclatureDokumen13 halamanKode Internasional NomenclaturekianaBelum ada peringkat
- Jurnal BiologiDokumen24 halamanJurnal BiologiDwi R. SavitriBelum ada peringkat
- Kelompok 1 ICN PREAMBLE, DIvision 1, 2 (Chapter 1)Dokumen7 halamanKelompok 1 ICN PREAMBLE, DIvision 1, 2 (Chapter 1)oyyyBelum ada peringkat
- Tata Nama TumbuhanDokumen36 halamanTata Nama TumbuhanhamzahB SKMBelum ada peringkat
- PaleontologiDokumen134 halamanPaleontologiHaposan S SamosirBelum ada peringkat
- Modul 2.ciri Mahkluk Hidup Dan KlasifikasiDokumen25 halamanModul 2.ciri Mahkluk Hidup Dan KlasifikasiDita NurulBelum ada peringkat
- Sistum I TM 1-7Dokumen216 halamanSistum I TM 1-7SABRINA ABEALLYA AFTHONI 1Belum ada peringkat
- Taksonomi Dan NomenklaturDokumen4 halamanTaksonomi Dan NomenklaturMoyo AdiBelum ada peringkat
- Zooin NewDokumen313 halamanZooin NewDeis Sesilia PanigoroBelum ada peringkat
- Evolusi MolekulerDokumen32 halamanEvolusi MolekulerSafaruddin Al AfganiBelum ada peringkat
- Kelompok 8 Tugas Resume Keanekaragaman Dan KlasifikasiDokumen32 halamanKelompok 8 Tugas Resume Keanekaragaman Dan Klasifikasisanti ariani rambeBelum ada peringkat
- Review KlasifikasiDokumen22 halamanReview KlasifikasianziaBelum ada peringkat
- Tatanama Tumbuhan Dan TanamanDokumen15 halamanTatanama Tumbuhan Dan TanamanFitri Utami Hasan100% (1)
- Resume Kelompok 4 - Keanekaragaman Dan Klasifikasi SeranggaDokumen21 halamanResume Kelompok 4 - Keanekaragaman Dan Klasifikasi SeranggaVira Putri FadhilahBelum ada peringkat
- Halaman 1Dokumen16 halamanHalaman 1Lailatul BadriyahBelum ada peringkat
- Makalah Materi 5 MK Phanerogamae - Camel G. MundaengDokumen17 halamanMakalah Materi 5 MK Phanerogamae - Camel G. MundaengCamel Gantonio MundaengBelum ada peringkat
- Summary - BAB 10 - Classification of MicroorganismDokumen11 halamanSummary - BAB 10 - Classification of MicroorganismGladys ElvinaBelum ada peringkat
- 1 PendahuluanDokumen18 halaman1 PendahuluanannisakarussaBelum ada peringkat
- BOTANI FARMASI Taksonomi Tumbuhan Diska NabillahDokumen5 halamanBOTANI FARMASI Taksonomi Tumbuhan Diska NabillahDiska Nabillah100% (1)
- Tatanama Mikrobia EDITDokumen31 halamanTatanama Mikrobia EDITSela AlifiaBelum ada peringkat
- Sejarah Perkembangan Taksonomi TumbuhanDokumen16 halamanSejarah Perkembangan Taksonomi TumbuhanAaaamaBelum ada peringkat
- Materi 7. Nomenklatur TumbuhanDokumen12 halamanMateri 7. Nomenklatur TumbuhanNanta MulyaBelum ada peringkat
- Laporan Hasil Diskusi Tugas 3 Kelompok 1Dokumen4 halamanLaporan Hasil Diskusi Tugas 3 Kelompok 1Bella Ayuana PutriBelum ada peringkat
- Kladogram (Biologi)Dokumen7 halamanKladogram (Biologi)fenny suhartiwiBelum ada peringkat
- Makalah Taksonomi Dan Sistematika Tumbuhan Dan Tugas Terstruktur 1Dokumen7 halamanMakalah Taksonomi Dan Sistematika Tumbuhan Dan Tugas Terstruktur 1Aku Big Bos DayuBelum ada peringkat
- Rangka Acara IiiDokumen27 halamanRangka Acara IiiSukma KinasihBelum ada peringkat
- Makalah Tata Nama TumbuhanDokumen8 halamanMakalah Tata Nama TumbuhanRahmat Dika JuniantoBelum ada peringkat
- Aspergillus NigerDokumen14 halamanAspergillus NigerSiti RositahBelum ada peringkat
- Makalah Klasifikasi Bakteri 10 OrdoDokumen12 halamanMakalah Klasifikasi Bakteri 10 OrdoIkhwan LatifBelum ada peringkat
- Botani Tumbuhan RendahDokumen18 halamanBotani Tumbuhan RendahRaodatul JannahBelum ada peringkat
- Makalah BTTDokumen17 halamanMakalah BTTEndoIeBelum ada peringkat
- EMBRIOLOGI AVES BLM FixDokumen21 halamanEMBRIOLOGI AVES BLM FixPatimmBelum ada peringkat
- SPH 2Dokumen22 halamanSPH 2PatimmBelum ada peringkat
- Poliembrio PDFDokumen15 halamanPoliembrio PDFPatimmBelum ada peringkat
- Makalah Inovasi Nata de SemangkaDokumen10 halamanMakalah Inovasi Nata de SemangkaPatimm100% (1)
- B - 1 - Sensasi InderaDokumen35 halamanB - 1 - Sensasi InderaPatimmBelum ada peringkat
- Penyajian DataDokumen32 halamanPenyajian DataPatimmBelum ada peringkat
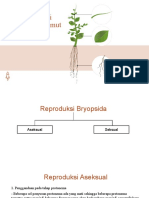
- Reproduksi LumutDokumen23 halamanReproduksi LumutPatimmBelum ada peringkat
- CHLORELLADokumen2 halamanCHLORELLAPatimmBelum ada peringkat
- Protista Mirip Jamur MyxomycotaDokumen10 halamanProtista Mirip Jamur MyxomycotaPatimmBelum ada peringkat
- Sistem Pencernaan SPH PDFDokumen34 halamanSistem Pencernaan SPH PDFPatimmBelum ada peringkat
- SynuraDokumen2 halamanSynuraPatimm100% (1)
- Universitas Negeri MalangDokumen14 halamanUniversitas Negeri MalangPatimmBelum ada peringkat
- Analisis Jurnal Biosel 1Dokumen3 halamanAnalisis Jurnal Biosel 1PatimmBelum ada peringkat