Anda mungkin juga menyukai
- RPPH TK-A SMT2 Minggu - 02 - Perlengkapan RekreasiDokumen6 halamanRPPH TK-A SMT2 Minggu - 02 - Perlengkapan RekreasiSri Maharaja Sanggrama Wijaya100% (5)
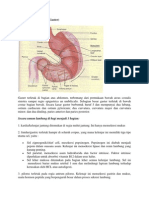
- Anatomi LambungDokumen7 halamanAnatomi LambungAtrioventrikular MilanistiBelum ada peringkat
- S1 Ars - Term PrefixesDokumen13 halamanS1 Ars - Term PrefixesCindy TifanaBelum ada peringkat
- Lamaran Pekerjaan Perawat di RSUD Dr. H. Chasan BoesorieDokumen1 halamanLamaran Pekerjaan Perawat di RSUD Dr. H. Chasan BoesorieSiti KartinaBelum ada peringkat
- Resep Obat PenyakitDokumen8 halamanResep Obat Penyakitmuliadewanti14Belum ada peringkat
- Referat Keracunan Asam KuatDokumen25 halamanReferat Keracunan Asam Kuatmamal malikaBelum ada peringkat
- RESEP PENULISANDokumen24 halamanRESEP PENULISANKartika RosyahBelum ada peringkat
- Soal Ujian Blok Aml CML FK - UnpriDokumen3 halamanSoal Ujian Blok Aml CML FK - UnpriAndy F MonroeBelum ada peringkat
- Anatomi 3 Dan 4Dokumen2 halamanAnatomi 3 Dan 4Arum Maharani LantagiBelum ada peringkat
- ProtistaDokumen1 halamanProtistaAyaBelum ada peringkat
- Apoptosis Dan NekrosisDokumen7 halamanApoptosis Dan NekrosisNaditya Fitriani HasanahBelum ada peringkat
- HistologiDokumen3 halamanHistologiNur akilaBelum ada peringkat
- Cara MembacaDokumen19 halamanCara MembacaIda Ayu PurnamaBelum ada peringkat
- Makalah Obat Wajib ApotekDokumen19 halamanMakalah Obat Wajib ApotekElsa ElsaBelum ada peringkat
- Oksidan & AntioksidanDokumen30 halamanOksidan & AntioksidanAdi Suryadi PutraBelum ada peringkat
- BEDAH MINOR DAN LUMBAL PUNCTIONDokumen7 halamanBEDAH MINOR DAN LUMBAL PUNCTIONTheofany SiraitBelum ada peringkat
- Sintesis ProteinDokumen39 halamanSintesis ProteinPutuBelum ada peringkat
- RESEP OBATDokumen28 halamanRESEP OBATAndriyani YaniBelum ada peringkat
- Presentasi BatukDokumen18 halamanPresentasi Batukjadi gini, nah gituBelum ada peringkat
- Terapi Obat Vasospasme Serebral Setelah Perdarahan Subaraknoid Akibat AneurismaDokumen14 halamanTerapi Obat Vasospasme Serebral Setelah Perdarahan Subaraknoid Akibat AneurismaRadi PdBelum ada peringkat
- Asma Patofisiologi dan Mekanisme ObatDokumen6 halamanAsma Patofisiologi dan Mekanisme ObatyunikBelum ada peringkat
- Soal Ujian Blok Panca Indera 2015Dokumen17 halamanSoal Ujian Blok Panca Indera 2015tanBelum ada peringkat
- LKK 4 ANAMNESIS THT - Sakit TenggorokanDokumen1 halamanLKK 4 ANAMNESIS THT - Sakit TenggorokanMuhammad AdityaBelum ada peringkat
- KELAINAN KELOPAK MATADokumen16 halamanKELAINAN KELOPAK MATAPasca RiandyBelum ada peringkat
- Elektrokardiografi: Dr. Surya Hafidiansyah Putra, SP - JP (K), Fiha, Fascc, FapscDokumen51 halamanElektrokardiografi: Dr. Surya Hafidiansyah Putra, SP - JP (K), Fiha, Fascc, FapscGiovani PurbaBelum ada peringkat
- Rigo Ginting 320200301019 AscomycotaDokumen5 halamanRigo Ginting 320200301019 AscomycotaGuardi FrandBelum ada peringkat
- Isian Borang Standar 4Dokumen21 halamanIsian Borang Standar 4Noval NovallBelum ada peringkat
- Analisis DNA ForensikDokumen31 halamanAnalisis DNA ForensikintanchairunnisaBelum ada peringkat
- InfeksiDokumen29 halamanInfeksiSarahBelum ada peringkat
- Taenia Solium Dan Taenia Saginata (Terjemahan)Dokumen10 halamanTaenia Solium Dan Taenia Saginata (Terjemahan)adhiBelum ada peringkat
- OBATANTEPRESANDokumen4 halamanOBATANTEPRESANIrfan SetyawanBelum ada peringkat
- ResepDokumen56 halamanResepTanani 102014007Belum ada peringkat
- MEDICAL TERMINOLOGY ROOTSDokumen23 halamanMEDICAL TERMINOLOGY ROOTSAnonymous DA8iQzBelum ada peringkat
- Resep UPKDokumen4 halamanResep UPKShendi SuryanaBelum ada peringkat
- Terminologi Medis ARSDokumen4 halamanTerminologi Medis ARSniaBelum ada peringkat
- B01. Arya Adhi Yoga Wikrama Jaya (018.06.0031) Laporan Praktikum Histologi Jaringan DasarDokumen29 halamanB01. Arya Adhi Yoga Wikrama Jaya (018.06.0031) Laporan Praktikum Histologi Jaringan DasarWJ AjaBelum ada peringkat
- Proses Ekskresi Urine dan Bersihan PlasmaDokumen8 halamanProses Ekskresi Urine dan Bersihan PlasmaingeBelum ada peringkat
- Mekanisme Kerja RifampisinDokumen8 halamanMekanisme Kerja RifampisinAfifah RizqiBelum ada peringkat
- Scrub Up and Gloving TecniqueDokumen6 halamanScrub Up and Gloving TecniqueBagus SamudroBelum ada peringkat
- Cara Kerja Lidah ManusiaDokumen2 halamanCara Kerja Lidah ManusiaIsman Bôèdâk SôëndàBelum ada peringkat
- Anfis HeparDokumen28 halamanAnfis HeparSolagrasiaBelum ada peringkat
- Matriks TulangDokumen3 halamanMatriks TulangSlamet IriyantoBelum ada peringkat
- Obat Anti JamurDokumen18 halamanObat Anti Jamurdhea permataBelum ada peringkat
- Cedera Dan Kematian SelularDokumen10 halamanCedera Dan Kematian SelularRitma WindartiBelum ada peringkat
- Farmakologi AntiaritmiaDokumen40 halamanFarmakologi AntiaritmiaKeton RcorBelum ada peringkat
- Makalah Penulngan EndokondralDokumen19 halamanMakalah Penulngan EndokondralAgako BerangkasBelum ada peringkat
- Diagnosis Dan Tatalaksana Penyakit Cacing Tambang: Hasil Anamnesis (Subjective)Dokumen3 halamanDiagnosis Dan Tatalaksana Penyakit Cacing Tambang: Hasil Anamnesis (Subjective)Edy KioonkBelum ada peringkat
- DNA DAN PROTEINDokumen6 halamanDNA DAN PROTEINYoga Pratama WijayaBelum ada peringkat
- Proses Menelan Secara MekanisDokumen3 halamanProses Menelan Secara MekanisNaylaBerlianaNugrahandhiniBelum ada peringkat
- SELA FUNGSIDokumen57 halamanSELA FUNGSIAsyifa Tsulus SandyBelum ada peringkat
- ASAM NUKLEAT DAN DNA EKSTRAKROMOSOMALDokumen37 halamanASAM NUKLEAT DAN DNA EKSTRAKROMOSOMALmutiara putri rahmawati0% (1)
- Resep OcaaDokumen10 halamanResep OcaaAZZHAARA RVBelum ada peringkat
- NeoplasmaDokumen48 halamanNeoplasmaNovika GabrielBelum ada peringkat
- Tahapan Moderator Pembukaan UmumDokumen8 halamanTahapan Moderator Pembukaan UmumOsissmk AlfabirBelum ada peringkat
- Disosiasi Dan IonisasiDokumen2 halamanDisosiasi Dan IonisasiKurrotiA'yunBelum ada peringkat
- Daya AmilolitisDokumen8 halamanDaya AmilolitisAnnisa IstiqamahBelum ada peringkat
- Macam MediaDokumen12 halamanMacam MediaRiaAffBelum ada peringkat
- Difusi Dan Dissolusi PDFDokumen16 halamanDifusi Dan Dissolusi PDFuwi nifahBelum ada peringkat
- Daftar Nama 144 Penyakit Yang Dapat Ditangani Di Layanan PrimerDokumen4 halamanDaftar Nama 144 Penyakit Yang Dapat Ditangani Di Layanan PrimerAbdul Anas100% (1)
- Mindmap Asam UratDokumen1 halamanMindmap Asam Uratpatrick star0% (1)
- Makalah Biologi Sel Kelompok 9 - Sintetis DnaDokumen16 halamanMakalah Biologi Sel Kelompok 9 - Sintetis DnaEvelliaBelum ada peringkat
- Makna Indonesia Sebagai Negara Hukum dan Ciri-Ciri Negara HukumDokumen6 halamanMakna Indonesia Sebagai Negara Hukum dan Ciri-Ciri Negara HukumNazwa DesfaniBelum ada peringkat
- LAPORAN TUGAS MANDIRI DASARDokumen42 halamanLAPORAN TUGAS MANDIRI DASARNazwa Desfani100% (2)
- 631 - Surat Progres Pengisian PPA Sampel Acak - Direktorat PAUDDokumen2 halaman631 - Surat Progres Pengisian PPA Sampel Acak - Direktorat PAUDNazwa DesfaniBelum ada peringkat
- Buku SDIDTK Bab I-VDokumen59 halamanBuku SDIDTK Bab I-VRahmatiaMozaike100% (1)
- Tema Dan Sub TemaDokumen5 halamanTema Dan Sub TemaismiaBelum ada peringkat
- Belajar Membaca Jilid 1Dokumen21 halamanBelajar Membaca Jilid 1Nazwa DesfaniBelum ada peringkat
- Modul Kuliah Bhs. Arab Pertemuan 7Dokumen5 halamanModul Kuliah Bhs. Arab Pertemuan 7Nazwa DesfaniBelum ada peringkat
- LK Hari Besar Nasional, Pahlawan NasionalDokumen9 halamanLK Hari Besar Nasional, Pahlawan NasionalNazwa DesfaniBelum ada peringkat
- SOP BAHASA INGGRISDokumen3 halamanSOP BAHASA INGGRISNazwa DesfaniBelum ada peringkat
- Modul Kuliah Bhs. Arab Pertemuan 4Dokumen5 halamanModul Kuliah Bhs. Arab Pertemuan 4Nazwa DesfaniBelum ada peringkat
- Modul Kuliah Bhs. Arab Pertemuan 5Dokumen5 halamanModul Kuliah Bhs. Arab Pertemuan 5Nazwa DesfaniBelum ada peringkat
- Akreditasi PAUD PNF Lampung 2021Dokumen19 halamanAkreditasi PAUD PNF Lampung 2021Nazwa DesfaniBelum ada peringkat
- LK Kendaraan AirDokumen9 halamanLK Kendaraan AirNeng Unie ZunetBelum ada peringkat
- Form Mengidentifikasi POTENSI Anak Dengan Kebutuhan KhususDokumen1 halamanForm Mengidentifikasi POTENSI Anak Dengan Kebutuhan KhususNazwa DesfaniBelum ada peringkat
- Und PesertaDokumen2 halamanUnd PesertaNazwa DesfaniBelum ada peringkat
- Panduan Registrasi Akun SipendiDokumen4 halamanPanduan Registrasi Akun SipendiNazwa DesfaniBelum ada peringkat
- Laporan SddidtkDokumen3 halamanLaporan SddidtkNazwa DesfaniBelum ada peringkat