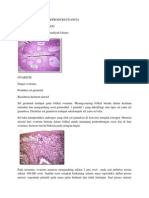
Anda mungkin juga menyukai
- Benang Dan Jarum BedahDokumen8 halamanBenang Dan Jarum BedahDessy Supitra Amiyati100% (6)
- ALARM ForcepsDokumen39 halamanALARM ForcepsCordova Arridho100% (1)
- 5 - Siklus MenstruasiDokumen46 halaman5 - Siklus MenstruasiIful SaifullahBelum ada peringkat
- Sperrof Lengkap 2 247 337Dokumen91 halamanSperrof Lengkap 2 247 337DESI SCORPINASARIBelum ada peringkat
- Siklus MenstruasiDokumen24 halamanSiklus MenstruasianggecintadiaBelum ada peringkat
- Siklus Ovarium dan ImplantasiDokumen9 halamanSiklus Ovarium dan Implantasidel Ajjah100% (1)
- AfcDokumen24 halamanAfcSeverina AdellaBelum ada peringkat
- PUADokumen41 halamanPUAAisyah AmirahBelum ada peringkat
- REPRODUKSIDokumen13 halamanREPRODUKSIZaki Ridhoni 2208Belum ada peringkat
- Fase LutealDokumen9 halamanFase LutealDeianeira ZhafirahBelum ada peringkat
- Fisiologi Siklus MenstruasiDokumen15 halamanFisiologi Siklus MenstruasiVertyBelum ada peringkat
- Dessy Elvira 1520332001Dokumen19 halamanDessy Elvira 1520332001Timmy LarasatiBelum ada peringkat
- Fisiologi HaidDokumen6 halamanFisiologi Haidaini venusBelum ada peringkat
- Sindrom Ovarium Polikistik REVISIDokumen30 halamanSindrom Ovarium Polikistik REVISIFitriya SyaifuddinBelum ada peringkat
- Regulasi Siklus Menstruasi #1Dokumen34 halamanRegulasi Siklus Menstruasi #1Dian ZilfiraBelum ada peringkat
- Sistem Reproduksi Pria dan WanitaDokumen40 halamanSistem Reproduksi Pria dan WanitaAnry Umar100% (3)
- Referat Teori 2 SelDokumen18 halamanReferat Teori 2 SelgungdeBelum ada peringkat
- Fisiologi Menstruasi NormalDokumen19 halamanFisiologi Menstruasi NormalHensBelum ada peringkat
- Fisiologi Menstruasi NormalDokumen10 halamanFisiologi Menstruasi NormalIka Ristianingrum100% (1)
- PBL 3 EndokrinDokumen28 halamanPBL 3 EndokrinRidho HidayatullohBelum ada peringkat
- SilverthornDokumen15 halamanSilverthornelizabeth tanBelum ada peringkat
- HAID TIDAK TERATURDokumen55 halamanHAID TIDAK TERATURRisna Wati100% (1)
- HORMON DAN SIKLUS MENSTRUASIDokumen4 halamanHORMON DAN SIKLUS MENSTRUASIputriimahkotaBelum ada peringkat
- Bahan MakalahDokumen20 halamanBahan MakalahLili HapverBelum ada peringkat
- Perubahan Endomerium Dalam Siklus Menstruasi PDFDokumen21 halamanPerubahan Endomerium Dalam Siklus Menstruasi PDFagus mardiyantoBelum ada peringkat
- Makalah FixDokumen16 halamanMakalah FixNurul IzzahBelum ada peringkat
- Resume Siklus Menstruasi - Rossy Aryanto Finanda - 22632110Dokumen4 halamanResume Siklus Menstruasi - Rossy Aryanto Finanda - 22632110Rossy Aryanto fBelum ada peringkat
- SherwoodDokumen13 halamanSherwoodelizabeth tanBelum ada peringkat
- Siklus MenstruasiDokumen8 halamanSiklus MenstruasiRia IdaparaBelum ada peringkat
- Siklus Hidup OvariumDokumen20 halamanSiklus Hidup OvariumHasanalitaBelum ada peringkat
- DUBDokumen46 halamanDUBedoBelum ada peringkat
- Fisiologi MenstruasiDokumen26 halamanFisiologi MenstruasiAnonymous pL7Ty4TVqBelum ada peringkat
- PERDARAHAN UTERUSDokumen33 halamanPERDARAHAN UTERUSAndi Arsyi AdlinaBelum ada peringkat
- Proses OvulasiDokumen15 halamanProses OvulasiJunaida Rahmi50% (2)
- Embriologi Dan TeratologiDokumen11 halamanEmbriologi Dan TeratologidrnyolBelum ada peringkat
- LAPORAN PBL Klmpok 7Dokumen16 halamanLAPORAN PBL Klmpok 7Ahmad MustakimBelum ada peringkat
- Capter 3 Implantasi, Embriogenesis, Dan Perkembangan Plasenta PDFDokumen38 halamanCapter 3 Implantasi, Embriogenesis, Dan Perkembangan Plasenta PDFTia Amalia Puti RenaeryBelum ada peringkat
- TEORI INSESSANT OVULATIONDokumen7 halamanTEORI INSESSANT OVULATIONHidayat BazeherBelum ada peringkat
- Fisiologi Reproduksi WanitaDokumen5 halamanFisiologi Reproduksi WanitaEva Rosalina100% (1)
- ANATOMIDANFUNGSIDINDINGUTERUSDokumen32 halamanANATOMIDANFUNGSIDINDINGUTERUSSasa AmelBelum ada peringkat
- Haid BerkelanjutanDokumen33 halamanHaid BerkelanjutanRJ7Belum ada peringkat
- Implantasi PlasentaDokumen14 halamanImplantasi PlasentaAdiesty adelliaBelum ada peringkat
- Bahan Ajar Histologi Reproduksi WanitaDokumen13 halamanBahan Ajar Histologi Reproduksi WanitaNovia RizqiBelum ada peringkat
- Menstruasi (Obstetri)Dokumen25 halamanMenstruasi (Obstetri)anisa putri utamiBelum ada peringkat
- Sperm, Folikulo, OogDokumen11 halamanSperm, Folikulo, OogFitria Ella WardaniBelum ada peringkat
- Materi Pertemuan 4Dokumen17 halamanMateri Pertemuan 4DD OBelum ada peringkat
- Oogenesis in Teleost FishDokumen7 halamanOogenesis in Teleost FishHillery Supith Pala'langanBelum ada peringkat
- Anatomi Kel 4Dokumen8 halamanAnatomi Kel 4shella arrumBelum ada peringkat
- Tugas Fisiologi Reproduksi Perempuan - Suratmi - 6411420028Dokumen6 halamanTugas Fisiologi Reproduksi Perempuan - Suratmi - 6411420028Suratmi UnnesBelum ada peringkat
- Arzia Sken5 - Fisiologi Siklus MenstruasiDokumen8 halamanArzia Sken5 - Fisiologi Siklus MenstruasiArzia RahmanBelum ada peringkat
- Mekanisme MenstruasiDokumen28 halamanMekanisme MenstruasiHendra Brahmantyo Ratsmananda100% (1)
- Perdarahan Uterus AbnormalDokumen50 halamanPerdarahan Uterus AbnormalMerlyn Chrislia RumtheBelum ada peringkat
- Perkembangan Minggu PertamaDokumen15 halamanPerkembangan Minggu Pertamahazia awanisBelum ada peringkat
- Mikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaDari EverandMikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaPenilaian: 2.5 dari 5 bintang2.5/5 (2)
- PSIKOLOGI PERKEMBANGAN ANAK DAN DISTURBILITAS PADA USIA EVOLUTIF: Apa itu dan bagaimana cara kerjanyaDari EverandPSIKOLOGI PERKEMBANGAN ANAK DAN DISTURBILITAS PADA USIA EVOLUTIF: Apa itu dan bagaimana cara kerjanyaBelum ada peringkat
- Mikrobiologi Medis I: Patogen dan Mikrobioma ManusiaDari EverandMikrobiologi Medis I: Patogen dan Mikrobioma ManusiaPenilaian: 4 dari 5 bintang4/5 (11)
- PSIKOLOGI, DEPRESI DAN DISTURBILITAS HUMOR: Memahami mekanisme dasarDari EverandPSIKOLOGI, DEPRESI DAN DISTURBILITAS HUMOR: Memahami mekanisme dasarPenilaian: 4 dari 5 bintang4/5 (3)
- Kegawatdaruratan Rujukan Obstetri (Dikes)Dokumen21 halamanKegawatdaruratan Rujukan Obstetri (Dikes)rake sardevaBelum ada peringkat
- Cara Kerja Beberapa Alat KontrasepsiDokumen11 halamanCara Kerja Beberapa Alat Kontrasepsirake sardevaBelum ada peringkat
- Kegawat Daruratan Di Bidang Obstetri (SG)Dokumen24 halamanKegawat Daruratan Di Bidang Obstetri (SG)rake sardevaBelum ada peringkat
- 00 Gu Pelatihan Akdrpp 251113Dokumen11 halaman00 Gu Pelatihan Akdrpp 251113rake sardevaBelum ada peringkat
- Op Tek MOW: Serial OptekDokumen27 halamanOp Tek MOW: Serial OptektapayanaBelum ada peringkat
- Alur Permohonan Rekomendasi STR 2020 DenpasarDokumen2 halamanAlur Permohonan Rekomendasi STR 2020 Denpasarrake sardevaBelum ada peringkat
- Asipirin Pada PreeklampsiaDokumen2 halamanAsipirin Pada Preeklampsiarake sardevaBelum ada peringkat
- Aspirin PEDokumen19 halamanAspirin PErake sardevaBelum ada peringkat
- Aspirin PEDokumen19 halamanAspirin PErake sardevaBelum ada peringkat
- Aspirin PEDokumen19 halamanAspirin PErake sardevaBelum ada peringkat
- 05 Ekstraks Vakum Forseps (Persalinan Perbantuan Alat - DRDokumen39 halaman05 Ekstraks Vakum Forseps (Persalinan Perbantuan Alat - DRrake sardevaBelum ada peringkat
- Berita Acara Perbaikan UP 3 NTDokumen10 halamanBerita Acara Perbaikan UP 3 NTrake sardevaBelum ada peringkat
- Modul 11 Asuhan KelahiranDokumen110 halamanModul 11 Asuhan Kelahiranrake sardevaBelum ada peringkat
- Fix PANDUAN PRAKTEK KLINIS 2018Dokumen10 halamanFix PANDUAN PRAKTEK KLINIS 2018rake sardevaBelum ada peringkat
- Antiphospholipid SyndromeDokumen16 halamanAntiphospholipid SyndromeKabir MuhammadBelum ada peringkat
- Fix Panduan Praktek Klinis 2018Dokumen89 halamanFix Panduan Praktek Klinis 2018rake sardeva100% (1)
- PARTUS KASEP RevisiDokumen33 halamanPARTUS KASEP Revisirake sardevaBelum ada peringkat
- Distosia ANADokumen28 halamanDistosia ANArake sardevaBelum ada peringkat
- Protap TransfusiDokumen11 halamanProtap Transfusirake sardevaBelum ada peringkat