Anda mungkin juga menyukai
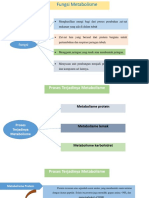
- Fungsi Dan Proses MetabolismeDokumen8 halamanFungsi Dan Proses MetabolismePutrik AgustinaBelum ada peringkat
- Cara KerjaOtotDokumen12 halamanCara KerjaOtotEu TyaBelum ada peringkat
- Modul 1 FistumDokumen12 halamanModul 1 FistumdefriantiBelum ada peringkat
- Biokimia Metabolisme Aerob Dan AnaerobDokumen4 halamanBiokimia Metabolisme Aerob Dan AnaerobarnalordBelum ada peringkat
- Integrasi MetabolismeDokumen3 halamanIntegrasi MetabolismeIcha PhynchaBelum ada peringkat
- Proses FesesDokumen3 halamanProses FesesElsya Meillinie100% (1)
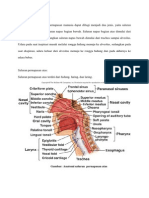
- Jaras Jaras PendengaranDokumen2 halamanJaras Jaras PendengaranSatriyani MuisBelum ada peringkat
- Rangkuman Metabolisme KarbohidratDokumen4 halamanRangkuman Metabolisme KarbohidratvutukhalBelum ada peringkat
- Eko SistemDokumen8 halamanEko SistemSarlotha TandiarrangBelum ada peringkat
- Metabolisme OtotDokumen2 halamanMetabolisme OtotDyahAyuRisqiNilamsariBelum ada peringkat
- Tugas Essay Fisiologi HewanDokumen4 halamanTugas Essay Fisiologi HewanKindi MuhamadBelum ada peringkat
- Struktur Mioglobin Dan HemoglobinDokumen5 halamanStruktur Mioglobin Dan HemoglobinShandyBelum ada peringkat
- Cara Kerja OtotDokumen64 halamanCara Kerja OtotSusanna ChanBelum ada peringkat
- Makalah Sistem Ekskresi ManusiaDokumen10 halamanMakalah Sistem Ekskresi ManusiaSalsabilla KusumaningrumBelum ada peringkat
- GlikolisisDokumen36 halamanGlikolisisAritias SimanjuntakBelum ada peringkat
- Biokimia-Ayu Puspitasari-Metabolisme Purin & Pirimidin 2014 PDFDokumen18 halamanBiokimia-Ayu Puspitasari-Metabolisme Purin & Pirimidin 2014 PDFAnonymous Uh9XtlGN100% (5)
- Tugas II Metabolisme KarbohidratDokumen5 halamanTugas II Metabolisme KarbohidratnuristhyqhBelum ada peringkat
- Sintesis Asam LaktatDokumen6 halamanSintesis Asam LaktatAhmad PratamaBelum ada peringkat
- Bio OptikDokumen49 halamanBio OptikNurhidayahBelum ada peringkat
- Biologi Uji MakananDokumen20 halamanBiologi Uji MakananGerry OxaBelum ada peringkat
- Mekanisme Kram OtotDokumen7 halamanMekanisme Kram OtotfitriBelum ada peringkat
- Metabolisme Di HatiDokumen24 halamanMetabolisme Di HaticresentiaBelum ada peringkat
- HMP ShuntDokumen12 halamanHMP ShuntDESY0% (1)
- Laporan Praktikum Uji Peragian Dan BenedictDokumen9 halamanLaporan Praktikum Uji Peragian Dan BenedictannisaBelum ada peringkat
- Makalah Kimia MipaDokumen12 halamanMakalah Kimia MipaFellan WestwoodBelum ada peringkat
- Proses Terjadinya Siklus OksigenDokumen3 halamanProses Terjadinya Siklus OksigenOsita PaboBelum ada peringkat
- Biokimia LipidDokumen25 halamanBiokimia LipidsovyrsBelum ada peringkat
- DEAMINASIDokumen13 halamanDEAMINASIseptiBelum ada peringkat
- Ekstremitas Superior (Edit)Dokumen17 halamanEkstremitas Superior (Edit)Priska Septri WePe100% (1)
- Organ Hati Pada ManusiaDokumen19 halamanOrgan Hati Pada Manusiaasdfghjkl22Belum ada peringkat
- Metabolisme NukleotidaDokumen13 halamanMetabolisme NukleotidaacelaBelum ada peringkat
- BIOLOGI SEL-1 (Lisosom Vakuola & Mikrobodi)Dokumen67 halamanBIOLOGI SEL-1 (Lisosom Vakuola & Mikrobodi)Elpe BibasBelum ada peringkat
- BAB III Pengaturan Osmolaritas CairanDokumen15 halamanBAB III Pengaturan Osmolaritas Cairanrara prameiBelum ada peringkat
- Fungsi Organ Sistem Penyusun Tubuh AvesDokumen8 halamanFungsi Organ Sistem Penyusun Tubuh AvesRobiatus SholikhahBelum ada peringkat
- Jurnal RespirasiDokumen4 halamanJurnal RespirasiMuhammad AsrizalBelum ada peringkat
- Gangguan Atau Penyakit Pada Sistem Pernapasan ManusiaDokumen6 halamanGangguan Atau Penyakit Pada Sistem Pernapasan Manusia082233665974Belum ada peringkat
- Sifat-Sifat Spektroskopik-Termal Asam Nukleat Kelompk 3 ADokumen10 halamanSifat-Sifat Spektroskopik-Termal Asam Nukleat Kelompk 3 ASerge WazukiBelum ada peringkat
- Makalah Biosel Kelompok 12 - Anabolisme KatabolismeDokumen17 halamanMakalah Biosel Kelompok 12 - Anabolisme KatabolismenurulBelum ada peringkat
- Perbedaan Respirasi Aerob Dan AnaerobDokumen2 halamanPerbedaan Respirasi Aerob Dan AnaerobTildisBelum ada peringkat
- Laporan Praktikum Faal 1 - Kelelahan OtotDokumen13 halamanLaporan Praktikum Faal 1 - Kelelahan OtotChanita ElonianTy100% (1)
- Glikogenesis, Glikogenolisis Dan GlukoneogenesisDokumen14 halamanGlikogenesis, Glikogenolisis Dan GlukoneogenesisAditya WillyBelum ada peringkat
- Proses Metabolisme Lemak Pada ManusiaDokumen6 halamanProses Metabolisme Lemak Pada ManusiaFance RuffBelum ada peringkat
- Infeksi Hati, Kantung Empedu, Dan PancreasDokumen41 halamanInfeksi Hati, Kantung Empedu, Dan PancreasdeviekaRDBelum ada peringkat
- Fenomena AuroraDokumen9 halamanFenomena AuroraeelBelum ada peringkat
- Katabolisme Rangka Karbon Dari Asam Amino BaruDokumen24 halamanKatabolisme Rangka Karbon Dari Asam Amino Barumimma amaliaBelum ada peringkat
- SIklus Asam Sitrat-1Dokumen19 halamanSIklus Asam Sitrat-1Salsa KhoirunisaBelum ada peringkat
- Laporan Sementara Cuka ApelDokumen18 halamanLaporan Sementara Cuka ApelRahma Diana YulistiahBelum ada peringkat
- Anatomi Saluran NapasDokumen5 halamanAnatomi Saluran NapasY3153NSBelum ada peringkat
- Buku BiokimiaDokumen150 halamanBuku BiokimiaNasroedien FikryBelum ada peringkat
- Jalur Asam Uronat VeDokumen8 halamanJalur Asam Uronat VesimplyveeBelum ada peringkat
- Oksidasi Beta Asam LemakDokumen2 halamanOksidasi Beta Asam LemakMeMed BoulalaBelum ada peringkat
- Katabolisme LipidDokumen30 halamanKatabolisme LipidAtiko Nur OktaVianiBelum ada peringkat
- Pengayaan Biokimia Kelompok 3 A2Dokumen58 halamanPengayaan Biokimia Kelompok 3 A2rullBelum ada peringkat
- Respirasi AerobDokumen13 halamanRespirasi AerobAnggii Mariisa Pase0% (1)
- Metabolisme Asam Amino & Protein, BASUKIHARDJO EDIT (H 24)Dokumen256 halamanMetabolisme Asam Amino & Protein, BASUKIHARDJO EDIT (H 24)Denok Ayu Mas Ruqoyyah100% (4)
- Dekarboksilasi Oksidatif & Siklus KrebsDokumen7 halamanDekarboksilasi Oksidatif & Siklus Krebsbella adisyaBelum ada peringkat
- Definisi Siklus KrebsDokumen9 halamanDefinisi Siklus Krebslamhot simamoraBelum ada peringkat
- TcaDokumen12 halamanTcaLala Latifah WiyandiBelum ada peringkat
- Siklus KrebsDokumen7 halamanSiklus KrebsDwi Arry NugraheniBelum ada peringkat
- Siklus Krep BiologiDokumen14 halamanSiklus Krep BiologiintanBelum ada peringkat
- Dokter Asing Vs Dokter Dalam NegriDokumen13 halamanDokter Asing Vs Dokter Dalam NegriSurya Ashri Rahman HBelum ada peringkat
- 13 LIMFADEMA DAN LIMFADENITIS DR - Dian ADokumen24 halaman13 LIMFADEMA DAN LIMFADENITIS DR - Dian ASurya Ashri Rahman HBelum ada peringkat
- Stress Manajemen Stress AnimasiDokumen23 halamanStress Manajemen Stress AnimasiNung TjahayaBelum ada peringkat
- Panduan Penulisan JurnalDokumen107 halamanPanduan Penulisan JurnalNarjus SafaahBelum ada peringkat
- Life Long LearningDokumen22 halamanLife Long LearningHenyAdriyantiRukmanaBelum ada peringkat
- 6 KELAINAN JANTUNG ANAK DR - TatangDokumen73 halaman6 KELAINAN JANTUNG ANAK DR - TatangSurya Ashri Rahman HBelum ada peringkat
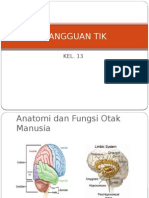
- Gangguan TikDokumen17 halamanGangguan TikShinta WulandhariBelum ada peringkat
- Daftar Pustaka NewDokumen16 halamanDaftar Pustaka NewSurya Ashri Rahman HBelum ada peringkat
- Konsep Dasar Pendidikan Kesehatan Masyarakat (PKM)Dokumen23 halamanKonsep Dasar Pendidikan Kesehatan Masyarakat (PKM)Narjus SafaahBelum ada peringkat
- Dasar-Dasar Komunikasi EfektifDokumen33 halamanDasar-Dasar Komunikasi EfektifNarjus SafaahBelum ada peringkat
- StreesDokumen10 halamanStreesakinariaBelum ada peringkat
- Infeksi Spesifik TGU PrintDokumen48 halamanInfeksi Spesifik TGU PrintSurya Ashri Rahman HBelum ada peringkat
- Membaca CepatDokumen18 halamanMembaca CepatSurya Ashri Rahman H100% (1)
- Gangguan TikDokumen16 halamanGangguan TikSurya Ashri Rahman HBelum ada peringkat
- Makala HDokumen28 halamanMakala HSurya Ashri Rahman HBelum ada peringkat
- Makala HDokumen7 halamanMakala HSurya Ashri Rahman HBelum ada peringkat
- Gangguan Kepribadian: SkizoidDokumen16 halamanGangguan Kepribadian: SkizoidSurya Ashri Rahman HBelum ada peringkat
- Makala HDokumen9 halamanMakala HSurya Ashri Rahman HBelum ada peringkat
- Makala HDokumen9 halamanMakala HSurya Ashri Rahman HBelum ada peringkat
- Makala HDokumen14 halamanMakala HSurya Ashri Rahman HBelum ada peringkat
- Makala HDokumen14 halamanMakala HSurya Ashri Rahman HBelum ada peringkat
- Makala HDokumen23 halamanMakala HSurya Ashri Rahman HBelum ada peringkat
- GDDokumen11 halamanGDSurya Ashri Rahman HBelum ada peringkat
- TR9 Autisme: HAR Omang RiskaDokumen23 halamanTR9 Autisme: HAR Omang RiskaSurya Ashri Rahman HBelum ada peringkat
- Makala HDokumen14 halamanMakala HSurya Ashri Rahman HBelum ada peringkat
- Hid Rose FalusDokumen21 halamanHid Rose FalusSurya Ashri Rahman HBelum ada peringkat
- Adenokarsinoma GinjalDokumen30 halamanAdenokarsinoma GinjalSurya Ashri Rahman HBelum ada peringkat
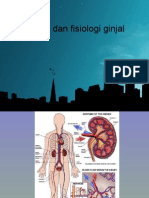
- Anatomi Fisiologi Ginjal111Dokumen27 halamanAnatomi Fisiologi Ginjal111Surya Ashri Rahman HBelum ada peringkat
- Abses Otak New JamakDokumen20 halamanAbses Otak New JamakSurya Ashri Rahman HBelum ada peringkat
- Bell's Palsy-Unizar - Mei 2015Dokumen29 halamanBell's Palsy-Unizar - Mei 2015Surya Ashri Rahman HBelum ada peringkat