Anda mungkin juga menyukai
- OPTIMASI TITRASIDokumen14 halamanOPTIMASI TITRASILaras Murni RahayuBelum ada peringkat
- Kimia Organik FisikDokumen27 halamanKimia Organik FisikIsmaMardaneniBelum ada peringkat
- Uji Disolusi CiproDokumen3 halamanUji Disolusi CiproHenri PrasetyoBelum ada peringkat
- Ekstraksi PelarutDokumen12 halamanEkstraksi PelarutGitha Jha DechBelum ada peringkat
- Per Mangano MetriDokumen9 halamanPer Mangano MetriLya WulandariBelum ada peringkat
- Percobaan1 Analisis ButanaDokumen8 halamanPercobaan1 Analisis ButanaZulvana Anggraeni HarvianBelum ada peringkat
- Kimanorfis Kel2Dokumen18 halamanKimanorfis Kel2Intan CahyaningrumBelum ada peringkat
- Aktinida Kelompok 3...Dokumen132 halamanAktinida Kelompok 3...Nova ArtawanBelum ada peringkat
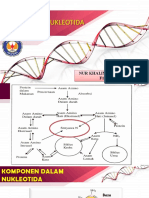
- Anabolisme NukleotidaDokumen17 halamanAnabolisme NukleotidaFirdhaa Blessing MvpBelum ada peringkat
- TemulawakAnalisisDokumen9 halamanTemulawakAnalisisanisa maryamBelum ada peringkat
- Kimia BioanorganikDokumen14 halamanKimia BioanorganikAgrina LismaBelum ada peringkat
- Faktor yang Mempengaruhi Aktivitas EnzimDokumen10 halamanFaktor yang Mempengaruhi Aktivitas EnzimDesyaniBelum ada peringkat
- Bab I MikrobiologiDokumen11 halamanBab I MikrobiologiWulan ClaxyBelum ada peringkat
- Alkohol Dehidrogenase Gaby MasyithaDokumen11 halamanAlkohol Dehidrogenase Gaby MasyithaLilik ZakiyaBelum ada peringkat
- Atom dan MolekulDokumen58 halamanAtom dan MolekulLelouch ViBelum ada peringkat
- Kromatografi Penukar IonDokumen10 halamanKromatografi Penukar IonImas WalijahBelum ada peringkat
- TEMBAGA UNSURDokumen6 halamanTEMBAGA UNSURfianBelum ada peringkat
- Percb 3 K1A021065 M. Dwi Jefry ArdiansyahDokumen19 halamanPercb 3 K1A021065 M. Dwi Jefry Ardiansyah28M. Dwi Jefry ArdiansyahBelum ada peringkat
- JUDULDokumen32 halamanJUDULRidho Arief Al RasyidBelum ada peringkat
- Pertemuan 1Dokumen13 halamanPertemuan 1Faradisa AninditaBelum ada peringkat
- Laporan 1 Kromatografi Penukar IonDokumen5 halamanLaporan 1 Kromatografi Penukar IonAirlangga Diandra Putra100% (1)
- REGULASI GENDokumen11 halamanREGULASI GENLivia Nur CholifahBelum ada peringkat
- DNA SEPENGENALANDokumen9 halamanDNA SEPENGENALANEka LuthfianaBelum ada peringkat
- UAS Kristalokimia - Salwa Kamilia - 15307141051Dokumen3 halamanUAS Kristalokimia - Salwa Kamilia - 15307141051Salwa KamiliaBelum ada peringkat
- Ekstraksi Pelarut OptimalDokumen9 halamanEkstraksi Pelarut OptimalShintaBelum ada peringkat
- ENZIM KARBOKSIPEPTIDASEDokumen13 halamanENZIM KARBOKSIPEPTIDASEFauziah Sulaeman D'nine ChastléBelum ada peringkat
- Skripsi KomplitDokumen75 halamanSkripsi KomplitErsan Yudhapratama Muslih100% (1)
- Model HuckelDokumen20 halamanModel HuckelAlfi FadhilaBelum ada peringkat
- Tugas 1 BiosintesisDokumen9 halamanTugas 1 BiosintesisTresnoningtias Mutiara AnisaBelum ada peringkat
- Teori Ikatan ValensiDokumen10 halamanTeori Ikatan ValensiBINTANGBelum ada peringkat
- Prediksi Jenis Ikatan Antara Kation Timbal (Ii) Dan Asam Humat Tinja Sapi (Ahts)Dokumen8 halamanPrediksi Jenis Ikatan Antara Kation Timbal (Ii) Dan Asam Humat Tinja Sapi (Ahts)Samik AmanahBelum ada peringkat
- MEKANISME REAKSI REDOKS INNER SPHEREDokumen2 halamanMEKANISME REAKSI REDOKS INNER SPHEREHaryadi AdityaBelum ada peringkat
- ElektroforesisDokumen13 halamanElektroforesisSteFunnie YolalandhaBelum ada peringkat
- V1/T1 = V2/T22/300 = 3/T2300×3 = 2×T2T2 = 450 K = 177 °CJadi, suhu gas tersebut adalah 177 °CDokumen23 halamanV1/T1 = V2/T22/300 = 3/T2300×3 = 2×T2T2 = 450 K = 177 °CJadi, suhu gas tersebut adalah 177 °CUrZeissBelum ada peringkat
- Presentasi Clock The Old Nassau Kelompok 11Dokumen22 halamanPresentasi Clock The Old Nassau Kelompok 11RimaBelum ada peringkat
- Katalis OrganologamDokumen10 halamanKatalis Organologamronawati silabanBelum ada peringkat
- NAHCO3 FUNGSIDokumen2 halamanNAHCO3 FUNGSIMaureen CelciliaBelum ada peringkat
- Proposal Kegiatan PmoDokumen8 halamanProposal Kegiatan PmoDewi MasulahBelum ada peringkat
- Lapres Asam AminoDokumen30 halamanLapres Asam AminoEka SeptianingtyasBelum ada peringkat
- Jurnal HepatotoksikDokumen6 halamanJurnal HepatotoksikDevy DjawaBelum ada peringkat
- Laporan Praktikum Percobaan 4Dokumen16 halamanLaporan Praktikum Percobaan 4Rizka Nur Yuniarsih100% (1)
- Jerry Febrico 2106706243 Isolasi DNADokumen11 halamanJerry Febrico 2106706243 Isolasi DNAJerry 689Belum ada peringkat
- Daun SirsakDokumen11 halamanDaun SirsakkikihandlerBelum ada peringkat
- Metode Titrasi ArgentometriDokumen1 halamanMetode Titrasi ArgentometriAdelia SetianingsihBelum ada peringkat
- REAKSI KARBOHIDRATDokumen20 halamanREAKSI KARBOHIDRATinnare' eBelum ada peringkat
- Laporan Praktikum Ekstraksi PelarutDokumen3 halamanLaporan Praktikum Ekstraksi PelarutFaizal AldinoBelum ada peringkat
- Laporan Praktikum Kimia Anorganik Uji ToksisitasDokumen8 halamanLaporan Praktikum Kimia Anorganik Uji ToksisitasRizky Widyastari100% (2)
- Laporan SpektrofotometriDokumen7 halamanLaporan SpektrofotometriyulinarBelum ada peringkat
- Clara Elisa - Laporan Modul02Dokumen11 halamanClara Elisa - Laporan Modul02Vicky Faras Barunson PanggabeanBelum ada peringkat
- Percobaan 7 - 062119058 - Raden Thasya Puteri. RDokumen9 halamanPercobaan 7 - 062119058 - Raden Thasya Puteri. RThasya PuteriBelum ada peringkat
- Sama Hap TCPDokumen5 halamanSama Hap TCPDewiBelum ada peringkat
- Tio 2Dokumen7 halamanTio 2Tommi0% (1)
- SN1 ReaksiDokumen43 halamanSN1 ReaksiSeliaDestianingrumBelum ada peringkat
- Sol Padat Intan HitamDokumen15 halamanSol Padat Intan HitamAbdurrofi RayadiBelum ada peringkat
- HUKUM LAMBERT BEERDokumen13 halamanHUKUM LAMBERT BEERSarah PanjaitanBelum ada peringkat
- 12-13.kromatografi 4, Penukar IonDokumen28 halaman12-13.kromatografi 4, Penukar IonAnny Saidatur100% (1)
- REGULASI ENZIMDokumen10 halamanREGULASI ENZIMEry RahmadBelum ada peringkat
- Laporan Praktikum FeSO4Dokumen8 halamanLaporan Praktikum FeSO4Amanda Putri100% (1)
- Siklus Asam SitratDokumen7 halamanSiklus Asam SitratSurya Ashri Rahman HBelum ada peringkat
- SKREBSINTESADokumen7 halamanSKREBSINTESADwi Arry NugraheniBelum ada peringkat
- KETENTUAN PRESENTASIDokumen2 halamanKETENTUAN PRESENTASIbella adisyaBelum ada peringkat
- Bab IV RokokDokumen2 halamanBab IV Rokokbella adisyaBelum ada peringkat
- Tinjauan Pustaka BHPDokumen10 halamanTinjauan Pustaka BHPbella adisyaBelum ada peringkat
- Anamnesis Penyakit JantungDokumen32 halamanAnamnesis Penyakit JantungMuhammad Fadli Balatif100% (4)
- Anemia Sel SabitDokumen3 halamanAnemia Sel Sabitbella adisyaBelum ada peringkat
- Bab I PKMDokumen5 halamanBab I PKMbella adisyaBelum ada peringkat
- Patofisiologi PsoriasisDokumen1 halamanPatofisiologi Psoriasisbella adisyaBelum ada peringkat
- Asam BasaDokumen8 halamanAsam Basabella adisyaBelum ada peringkat
- Anemia Sel SabitDokumen3 halamanAnemia Sel Sabitbella adisyaBelum ada peringkat
- Neuron NeurogliaDokumen3 halamanNeuron Neurogliabella adisyaBelum ada peringkat
- Farmako GlaukomaDokumen25 halamanFarmako Glaukomabella adisyaBelum ada peringkat
- CASE 2 KonjungtivitisDokumen5 halamanCASE 2 Konjungtivitisbella adisyaBelum ada peringkat
- Ulkus TungkaiDokumen34 halamanUlkus Tungkaibella adisyaBelum ada peringkat
- Para Thy PusDokumen6 halamanPara Thy Pusbella adisyaBelum ada peringkat
- TRANSPORTASI LIPID DALAM TUBUHDokumen5 halamanTRANSPORTASI LIPID DALAM TUBUHbella adisyaBelum ada peringkat
- Perawatan Pasien Luka BakarDokumen22 halamanPerawatan Pasien Luka Bakarbella adisyaBelum ada peringkat
- Dna Mito & IntiDokumen5 halamanDna Mito & Intibella adisyaBelum ada peringkat
- Faal KulitDokumen11 halamanFaal Kulitbella adisyaBelum ada peringkat
- Patofisiologi Dermatitis NumularisDokumen1 halamanPatofisiologi Dermatitis Numularisbella adisyaBelum ada peringkat
- Ulkus TungkaiDokumen34 halamanUlkus Tungkaibella adisyaBelum ada peringkat
- KISTA GANGLION: PENYAKIT YANG MENYEBABKAN BENJOLAN DI PERGELANGAN TANGANDokumen17 halamanKISTA GANGLION: PENYAKIT YANG MENYEBABKAN BENJOLAN DI PERGELANGAN TANGANbella adisyaBelum ada peringkat
- Bagaimana Sel Dapat Membentuk TubuhDokumen3 halamanBagaimana Sel Dapat Membentuk Tubuhbella adisyaBelum ada peringkat
- Bentuk Sediaan ObatDokumen28 halamanBentuk Sediaan Obatbella adisyaBelum ada peringkat
- Laporan Case NeoplasmaDokumen2 halamanLaporan Case Neoplasmabella adisyaBelum ada peringkat
- Cedera Otot, Tendon, LigamenDokumen30 halamanCedera Otot, Tendon, Ligamenbella adisyaBelum ada peringkat
- Buku Ilmu Penyakit Mata UgmDokumen375 halamanBuku Ilmu Penyakit Mata UgmGalih Yogo88% (17)
- Flipchart NeoplasmaDokumen1 halamanFlipchart Neoplasmabella adisyaBelum ada peringkat
- Asam BasaDokumen8 halamanAsam Basabella adisyaBelum ada peringkat
- Bentuk Sediaan ObatDokumen28 halamanBentuk Sediaan Obatbella adisyaBelum ada peringkat