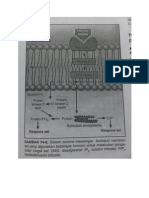
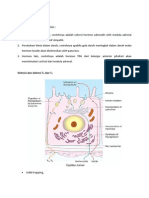
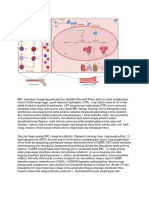
Anda mungkin juga menyukai
- 2021 - MKD101 - Sub-Topik 2. Jalur Transduksi Sinyal Hormon Melalui AMP Siklik Dan Second Messenger LainnyaDokumen2 halaman2021 - MKD101 - Sub-Topik 2. Jalur Transduksi Sinyal Hormon Melalui AMP Siklik Dan Second Messenger LainnyayanaBelum ada peringkat
- Oxytocin Dan FSHDokumen6 halamanOxytocin Dan FSHDwi agustinaBelum ada peringkat
- AdrenergikDokumen9 halamanAdrenergikRavi DasmanBelum ada peringkat
- Second MassengerDokumen7 halamanSecond MassengerMa MedBelum ada peringkat
- PROTEIN G Dan Second MessengerDokumen5 halamanPROTEIN G Dan Second Messengerelfira_handayaniBelum ada peringkat
- Mekanisme Komunikasi SelDokumen5 halamanMekanisme Komunikasi SelbabalkhanBelum ada peringkat
- Mekanisme Transduksi Sinyal KimiaDokumen4 halamanMekanisme Transduksi Sinyal KimiaHitaHusmarikaBelum ada peringkat
- Materi Presentasi Biosel TransduksiDokumen4 halamanMateri Presentasi Biosel TransduksiNurcahyatiBelum ada peringkat
- Modul Pertemuan 12 Siklus Asam SitratDokumen6 halamanModul Pertemuan 12 Siklus Asam SitratsempoErna naaBelum ada peringkat
- Jaawaban Contoh Soal UTSDokumen16 halamanJaawaban Contoh Soal UTSLinda Trivana HavanaBelum ada peringkat
- Jalur EMP Heksosa DifosfatDokumen47 halamanJalur EMP Heksosa DifosfatKahfi ArdhianBelum ada peringkat
- Metabolisme (Katabolisme Dan Anabolisme)Dokumen47 halamanMetabolisme (Katabolisme Dan Anabolisme)Auhge Eva Sari ManikBelum ada peringkat
- Peranan ATP Dalam Proses Metabolisme Beserta KelainannyaDokumen28 halamanPeranan ATP Dalam Proses Metabolisme Beserta KelainannyaarvindaBelum ada peringkat
- Rantik Biosinyal TBC 2021Dokumen44 halamanRantik Biosinyal TBC 2021Nurha LizaBelum ada peringkat
- MP 2Dokumen3 halamanMP 2bengbengBelum ada peringkat
- Regulasi MetabolismeDokumen16 halamanRegulasi MetabolismeHendrik_NurfitriantoBelum ada peringkat
- Penerimaan Sinyal Dan Inisiasi TransduksiDokumen8 halamanPenerimaan Sinyal Dan Inisiasi TransduksiVica FirmaBelum ada peringkat
- Proses Glikolisis Secara Singkat Dan PenjelasannyaDokumen5 halamanProses Glikolisis Secara Singkat Dan PenjelasannyaIzzah FarhanaBelum ada peringkat
- Proses Glikolisis Secara Singkat Dan PenjelasannyaDokumen5 halamanProses Glikolisis Secara Singkat Dan PenjelasannyaIzzah FarhanaBelum ada peringkat
- Mekanisme Kerja HormonDokumen2 halamanMekanisme Kerja HormonSaga SabaraBelum ada peringkat
- Translate Buku Kaplan&Sadok's Comprehensive Textbook Psychiaty 8th Edition. Hal 91-100Dokumen24 halamanTranslate Buku Kaplan&Sadok's Comprehensive Textbook Psychiaty 8th Edition. Hal 91-100Timothy M. PoluanBelum ada peringkat
- Tugas Baca FarmakoDokumen2 halamanTugas Baca FarmakoMazida maghfira putriBelum ada peringkat
- Mekanisme Ctm-GproteinDokumen9 halamanMekanisme Ctm-GproteinAdelia FryhaniBelum ada peringkat
- Kontrol Sekresi HormonDokumen10 halamanKontrol Sekresi HormonMuhamadRagaBelum ada peringkat
- Mekanisme Kerja HormonDokumen3 halamanMekanisme Kerja HormonRendi SiregarBelum ada peringkat
- Presentasi Biologi MolekulerDokumen21 halamanPresentasi Biologi Molekulerlieken mehingkoBelum ada peringkat
- Terjemahan CampbellDokumen3 halamanTerjemahan CampbellMirsa LeanderBelum ada peringkat
- Ujian Akhir SemesterDokumen18 halamanUjian Akhir SemesterfauziaBelum ada peringkat
- Pokbing IKD1 Case2Dokumen7 halamanPokbing IKD1 Case2Ressa DyanaBelum ada peringkat
- Biokimia Tugas3Dokumen4 halamanBiokimia Tugas3Ulfa Tri Mulyani NasutionBelum ada peringkat
- FarMol Transduksi Sinyal SelDokumen24 halamanFarMol Transduksi Sinyal SelRifqi UtomoBelum ada peringkat
- Metabolisme KarbohidratDokumen100 halamanMetabolisme KarbohidratIdank MunawarBelum ada peringkat
- PBL Blok11 Metabolik EndokrinDokumen30 halamanPBL Blok11 Metabolik EndokrinputuadiputraBelum ada peringkat
- BiokimiaDokumen4 halamanBiokimiaWindy Juwita Lumbanraja 20000002Belum ada peringkat
- Pelepasan Dan Sintesis HormonDokumen76 halamanPelepasan Dan Sintesis HormonKangjul AripinBelum ada peringkat
- Tugas Biologi MolekulerDokumen2 halamanTugas Biologi MolekulerMahda Rizki LianaBelum ada peringkat
- G Protein Coupled ReceptorDokumen11 halamanG Protein Coupled ReceptorFastabiqul KhairatBelum ada peringkat
- Klasifikasi HormonDokumen3 halamanKlasifikasi HormonRisa Maulida WidjayaBelum ada peringkat
- HORMONDokumen43 halamanHORMONSyamsul BachriBelum ada peringkat
- Metabolisme KarbohidratDokumen16 halamanMetabolisme KarbohidratSarni SarniBelum ada peringkat
- Transduksi SinyalDokumen78 halamanTransduksi SinyalArimbi Purinada MiandariBelum ada peringkat
- Materi 1 Sel Dan Organel SelDokumen34 halamanMateri 1 Sel Dan Organel SelsuambayogaBelum ada peringkat
- Jawaban No 2 3Dokumen2 halamanJawaban No 2 3yayansyaifan210804Belum ada peringkat
- Metabolisme Karbohidrat Ms WordDokumen18 halamanMetabolisme Karbohidrat Ms WordMutia DwisaBelum ada peringkat
- Metabolisme Tubuh (Glukosa)Dokumen26 halamanMetabolisme Tubuh (Glukosa)Arie FandyBelum ada peringkat
- Mekanisme Kerja REDokumen6 halamanMekanisme Kerja REkeylalakey1234Belum ada peringkat
- Materi Biologi Kelas 10, 11, 12Dokumen28 halamanMateri Biologi Kelas 10, 11, 12Rayyan MuzakyBelum ada peringkat
- Komunikasi Antar SelDokumen24 halamanKomunikasi Antar SelNunik Utari Nurwulandari100% (1)
- Metabolisme ProteinDokumen25 halamanMetabolisme ProteinHadyan TamimiBelum ada peringkat
- Rezwan Mulya Sipahutar - Biomet - Pskm19aDokumen3 halamanRezwan Mulya Sipahutar - Biomet - Pskm19ajuan sipaBelum ada peringkat
- Biokimia Karbohidrat & ProteinDokumen27 halamanBiokimia Karbohidrat & ProteinIvana ChristyBelum ada peringkat
- Protein G Farmakologi (Kel.4)Dokumen30 halamanProtein G Farmakologi (Kel.4)Mutia Rahayu100% (7)
- Learning Objective Skenario 4Dokumen3 halamanLearning Objective Skenario 4Lalu Wisnu ramadhanBelum ada peringkat
- Tugas 2 Biokimia Kel 2Dokumen8 halamanTugas 2 Biokimia Kel 2Elda AlfiyaniBelum ada peringkat
- MODUL BIOKIM Intergrasi MetabolismeDokumen18 halamanMODUL BIOKIM Intergrasi MetabolismeSiti AnisahBelum ada peringkat
- FarmakologiDokumen13 halamanFarmakologijihan febriyantiBelum ada peringkat
- Regulasi MetabolikDokumen34 halamanRegulasi MetabolikHesti NurrohmahBelum ada peringkat
- C.. Metabolisma (Revisi)Dokumen109 halamanC.. Metabolisma (Revisi)Syafira MpuhuBelum ada peringkat
- NukleolusDokumen2 halamanNukleolusSiska NurfauziaBelum ada peringkat
- KKO Taksonomi Bloom Versi Revisi BaruDokumen2 halamanKKO Taksonomi Bloom Versi Revisi BaruSiska NurfauziaBelum ada peringkat
- Peraturan Akademik MadrasahDokumen11 halamanPeraturan Akademik MadrasahSiska NurfauziaBelum ada peringkat
- SSDokumen24 halamanSSSiska NurfauziaBelum ada peringkat
- Contoh Surat Permohonan Pengesahan Dokumen I KTSP, Lembar Pengesahan, Rekomendasi, Dan InstrumenDokumen5 halamanContoh Surat Permohonan Pengesahan Dokumen I KTSP, Lembar Pengesahan, Rekomendasi, Dan InstrumenSiska NurfauziaBelum ada peringkat
- Kesimpulan Edm-DikonversiDokumen7 halamanKesimpulan Edm-DikonversisihabudinBelum ada peringkat
- Alumni Akper 2016Dokumen29 halamanAlumni Akper 2016Siska NurfauziaBelum ada peringkat
- No Urut LombaDokumen1 halamanNo Urut LombaSiska NurfauziaBelum ada peringkat
- Ashyatnya Manfaat Air Susu IbuDokumen2 halamanAshyatnya Manfaat Air Susu IbuSiska NurfauziaBelum ada peringkat
- Pembentukan Sel KelaminDokumen2 halamanPembentukan Sel KelaminSiska NurfauziaBelum ada peringkat
- JJJJJJDokumen14 halamanJJJJJJIka Wardhani KaruniaBelum ada peringkat
- Maklumat Kepulangan Santri Ramadhan 1442 H.-1Dokumen1 halamanMaklumat Kepulangan Santri Ramadhan 1442 H.-1Siska NurfauziaBelum ada peringkat
- RPP Sistem Reproduksi - Siska Nurfauzia P.Biologi 12Dokumen11 halamanRPP Sistem Reproduksi - Siska Nurfauzia P.Biologi 12Siska NurfauziaBelum ada peringkat
- Kurikulum Pemetaan Sistem ReproduksiDokumen5 halamanKurikulum Pemetaan Sistem ReproduksiSiska NurfauziaBelum ada peringkat
- RPP Tik Kelas 9Dokumen35 halamanRPP Tik Kelas 9Siska NurfauziaBelum ada peringkat
- Materi Cpns TKDDokumen62 halamanMateri Cpns TKDAlva Zalta100% (3)
- RK13AR10FIS0704Dokumen36 halamanRK13AR10FIS0704Devi SaidullohBelum ada peringkat
- MAPPING CURRICULUM BIOLOGI XI - Siska Nurfauzia P.Biologi'12Dokumen9 halamanMAPPING CURRICULUM BIOLOGI XI - Siska Nurfauzia P.Biologi'12Siska NurfauziaBelum ada peringkat
- Percobaan EkskresiDokumen5 halamanPercobaan EkskresiZakiyyatul MiskiyahBelum ada peringkat
- Percobaan EkskresiDokumen5 halamanPercobaan EkskresiZakiyyatul MiskiyahBelum ada peringkat
- LPK ProdukDokumen1 halamanLPK ProdukSiska NurfauziaBelum ada peringkat
- LPK ProyekDokumen2 halamanLPK ProyekJaeCk_GFourBelum ada peringkat
- LPK Unjuk KerjaDokumen2 halamanLPK Unjuk KerjaJaeCk_GFourBelum ada peringkat
- Lampiran Teknik Dan Instrumen PenilaianDokumen21 halamanLampiran Teknik Dan Instrumen Penilaiandairyplasma2163Belum ada peringkat
- Kalender Pendidikan 2017-2018Dokumen14 halamanKalender Pendidikan 2017-2018Mustari StarlyBelum ada peringkat
- LPK PortofolioDokumen1 halamanLPK PortofolioSiska NurfauziaBelum ada peringkat
- Kode Etik, 2. Ikrar Guru, 3. Tata Tertib Guru, 4. Alokasi Waktu, 5. Pembiasaan GuruDokumen23 halamanKode Etik, 2. Ikrar Guru, 3. Tata Tertib Guru, 4. Alokasi Waktu, 5. Pembiasaan Guruabah94% (31)
- Jurnal Guru MengajarDokumen5 halamanJurnal Guru MengajarFatriPopo100% (1)
- Kode Etik, 2. Ikrar Guru, 3. Tata Tertib Guru, 4. Alokasi Waktu, 5. Pembiasaan GuruDokumen23 halamanKode Etik, 2. Ikrar Guru, 3. Tata Tertib Guru, 4. Alokasi Waktu, 5. Pembiasaan Guruabah94% (31)