Anda mungkin juga menyukai
- Makalah DNA RekombinanDokumen9 halamanMakalah DNA RekombinanNisa TazkiyahBelum ada peringkat
- Jurnal FarmasiDokumen7 halamanJurnal FarmasiAry PratiwiBelum ada peringkat
- Enzim RestriksiDokumen3 halamanEnzim RestriksiSurya AdityaBelum ada peringkat
- Mikrobiologi TerapanDokumen3 halamanMikrobiologi TerapanSuhaila Zakiya NajahBelum ada peringkat
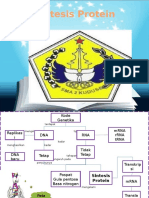
- Sintesis Protein Adalah Proses Di Mana Rantai Polipeptida Dibentuk Dari Kombinasi Kode Asam Amino Tunggal Di Dalam SelDokumen6 halamanSintesis Protein Adalah Proses Di Mana Rantai Polipeptida Dibentuk Dari Kombinasi Kode Asam Amino Tunggal Di Dalam SelWildanul HasanahBelum ada peringkat
- Pengertian ApoptosisDokumen23 halamanPengertian ApoptosisYunita Afina100% (1)
- Morfogenesis & Differensiasi - SelDokumen83 halamanMorfogenesis & Differensiasi - SelRizuka FathanahBelum ada peringkat
- Makalah MikrobiologiDokumen23 halamanMakalah MikrobiologiEvi Anna Tri SutrisnoBelum ada peringkat
- Prinsip Dan Pemanfaatan Rekayasa Genetik Dalam Produksi ObatDokumen40 halamanPrinsip Dan Pemanfaatan Rekayasa Genetik Dalam Produksi ObatFee ChocoBelum ada peringkat
- Vektor KloningDokumen10 halamanVektor KloningAgil Cendoll AnggaraBelum ada peringkat
- Sida RhombifoliaDokumen5 halamanSida RhombifoliaMelzy Putri SaniBelum ada peringkat
- Laporan - Praktikum Biologi Mikroba TropisDokumen18 halamanLaporan - Praktikum Biologi Mikroba Tropisbiowan2509Belum ada peringkat
- Kultur KalusDokumen35 halamanKultur KalusArmansyah maulana harahapBelum ada peringkat
- DNA BarcodingDokumen9 halamanDNA BarcodingRisma KareninaBelum ada peringkat
- Makalah Bukti EvolusiDokumen11 halamanMakalah Bukti EvolusiGrattyBelum ada peringkat
- 1Dokumen24 halaman1ArwindaBelum ada peringkat
- Metabolit Sekunder Poliamina Pada TumbuhanDokumen11 halamanMetabolit Sekunder Poliamina Pada TumbuhanHumaedii Hanya-NyaBelum ada peringkat
- Mitosis Akar BawangDokumen14 halamanMitosis Akar BawangArif Rachmatullah100% (1)
- Plasmid Dan EpisomDokumen8 halamanPlasmid Dan EpisomWidyarnes N. TikaBelum ada peringkat
- Laporan Praktikum Kuljar - Vicky Anggara - 061118043Dokumen9 halamanLaporan Praktikum Kuljar - Vicky Anggara - 061118043Vicky Anggara100% (1)
- One Gene One Polypeptide HypothesisDokumen8 halamanOne Gene One Polypeptide HypothesisErvicka CahyaBelum ada peringkat
- Apa Manfaat Rekayasa GenetikaDokumen8 halamanApa Manfaat Rekayasa GenetikaEpi NurafniBelum ada peringkat
- Makalah Kel 5 BioluminensiDokumen12 halamanMakalah Kel 5 BioluminensiPutri Aliyah100% (1)
- Bioteknologi PertanianDokumen10 halamanBioteknologi PertanianIRWANDI HUTBAH100% (1)
- Isolasi DnaDokumen11 halamanIsolasi DnaAnitaCitraAgustina0% (1)
- Sintesis ProteinDokumen29 halamanSintesis ProteinAtika RizkiBelum ada peringkat
- Soal Quiz Regulasi Ekspresi GenDokumen4 halamanSoal Quiz Regulasi Ekspresi GenElfrida MalauBelum ada peringkat
- Biokontrol Kapang Antagonis Terhadap Kapang PatogenDokumen4 halamanBiokontrol Kapang Antagonis Terhadap Kapang PatogenAlifia RahmawatiBelum ada peringkat
- Jurnal GENETIKA Siklus Hidup DrosophilaDokumen18 halamanJurnal GENETIKA Siklus Hidup DrosophilaSiti SoffahBelum ada peringkat
- Isolasi Dna N ElektroforesisDokumen23 halamanIsolasi Dna N ElektroforesisPrima Santi0% (1)
- Biomolekuler GenebankDokumen1 halamanBiomolekuler GenebankAmir CandraBelum ada peringkat
- Morfologi Virus Dna Dan RnaDokumen8 halamanMorfologi Virus Dna Dan RnaagnesliliantandjungBelum ada peringkat
- Resume Genetika KelaminDokumen9 halamanResume Genetika KelaminGufron AlifiBelum ada peringkat
- Makalah Transformasi SelDokumen40 halamanMakalah Transformasi Selanesco tuasuun0% (1)
- XIII-XIV Asimilasi NitrogenDokumen19 halamanXIII-XIV Asimilasi NitrogencHuAz TeeKaa0% (1)
- Laporan Praktikum Versi Mini RisetDokumen48 halamanLaporan Praktikum Versi Mini Risetnelmian100% (1)
- Artikel Mikroteknik Mengidentifikasi Isolasi Dna Pada BuahDokumen8 halamanArtikel Mikroteknik Mengidentifikasi Isolasi Dna Pada BuahPutri Mustika PratiwiBelum ada peringkat
- Resume Materi Genetika (Rekayasa Genetika)Dokumen2 halamanResume Materi Genetika (Rekayasa Genetika)fifahfifahBelum ada peringkat
- Sel Mikroba Dan StrukturnyaDokumen4 halamanSel Mikroba Dan StrukturnyaNur Rizqiatul Aulia100% (1)
- Hukum EvoluDokumen7 halamanHukum EvoluSitri SaripiBelum ada peringkat
- Fistum Kelompok 9 Topik HormonDokumen75 halamanFistum Kelompok 9 Topik HormonDiah Ayu PradnyaniBelum ada peringkat
- Replikasi Aparatus Phage t4Dokumen11 halamanReplikasi Aparatus Phage t4Friska WardahnyBelum ada peringkat
- Efisiensi Metabolisme Pada Hewan Vertebrata (AutDokumen9 halamanEfisiensi Metabolisme Pada Hewan Vertebrata (AutNisrina Setiowati100% (1)
- Laporan Proyek Sains Tumbuhan AgniaDokumen18 halamanLaporan Proyek Sains Tumbuhan AgniaAgniaVibrianiBelum ada peringkat
- Study Perilaku MonyetDokumen25 halamanStudy Perilaku MonyetSiti MutmainahBelum ada peringkat
- Kode Genetik ResumeDokumen12 halamanKode Genetik Resumeimroatun nafi'ahBelum ada peringkat
- Plasmid & EpisomDokumen13 halamanPlasmid & EpisomNandoBelum ada peringkat
- Ekspresi Gen Pada EukariotDokumen12 halamanEkspresi Gen Pada EukariotAssayid Muhammad Haqqi AnnaziliBelum ada peringkat
- Aplikasi Praktis Dari Teknologi DnaDokumen12 halamanAplikasi Praktis Dari Teknologi DnaGetta Austin MangiringBelum ada peringkat
- REVISI Kecenderungan Kawin Indeks Isolasi PDFDokumen38 halamanREVISI Kecenderungan Kawin Indeks Isolasi PDFDiah Ajeng MustikariniBelum ada peringkat
- Teori Evolusi Pasca DarwinDokumen4 halamanTeori Evolusi Pasca DarwinLivia Nur CholifahBelum ada peringkat
- Bab Vi Produksi Metabolit SekunderDokumen98 halamanBab Vi Produksi Metabolit SekunderGabriel AshariBelum ada peringkat
- Bioteknologi FermentasiDokumen9 halamanBioteknologi FermentasiNova Tri Kusuma DewiBelum ada peringkat
- Produksi Metabolit Sekunder Dengan Kultur JaringanDokumen17 halamanProduksi Metabolit Sekunder Dengan Kultur JaringanYulia Dyah SavitriBelum ada peringkat
- LAPORAN Induksi Tunas Kalus (BAB 1 & BAB 2)Dokumen20 halamanLAPORAN Induksi Tunas Kalus (BAB 1 & BAB 2)Hayyu ArumsariBelum ada peringkat
- Laporan Perbanyakan StarterDokumen11 halamanLaporan Perbanyakan StarterJane Jeng-jengBelum ada peringkat
- Makalah DNA Rekombinan 2Dokumen27 halamanMakalah DNA Rekombinan 2Ridwan NurdinBelum ada peringkat
- FiksDokumen23 halamanFiksMoeLkhaiva MundingBelum ada peringkat
- Biologi MolekulerDokumen9 halamanBiologi MolekulerEka LuthfianaBelum ada peringkat
- Kel.7 Rekayasa Genetika BiokimiamolekulerDokumen30 halamanKel.7 Rekayasa Genetika BiokimiamolekulerBang OtBelum ada peringkat
- Bioteknologi Molekul Kel.3Dokumen23 halamanBioteknologi Molekul Kel.3MoeLkhaiva MundingBelum ada peringkat
- 5 - 2120424781 - Wiwit Zuriati UnoDokumen1 halaman5 - 2120424781 - Wiwit Zuriati UnoWiwit Zuriati UnoBelum ada peringkat
- Wiwit Zuriati Uno 2120424781Dokumen4 halamanWiwit Zuriati Uno 2120424781Wiwit Zuriati UnoBelum ada peringkat
- 5 NIM NamaDokumen1 halaman5 NIM NamaANDI SETIAWANBelum ada peringkat
- 4 NIM NamaDokumen1 halaman4 NIM NamaWiwit Zuriati UnoBelum ada peringkat
- Wiwit Zuriati Uno - 2120424791 - CD-IO BudewiDokumen7 halamanWiwit Zuriati Uno - 2120424791 - CD-IO BudewiWiwit Zuriati UnoBelum ada peringkat
- Rangkuman Resep C2Dokumen10 halamanRangkuman Resep C2Wiwit Zuriati UnoBelum ada peringkat
- 3 NIM NamaDokumen1 halaman3 NIM NamaWiwit Zuriati UnoBelum ada peringkat
- Nim NamaDokumen4 halamanNim NamaWiwit Zuriati UnoBelum ada peringkat
- Bab II Tipus IntranasalDokumen25 halamanBab II Tipus IntranasalWiwit Zuriati UnoBelum ada peringkat
- Fish Bone c4Dokumen1 halamanFish Bone c4Wiwit Zuriati UnoBelum ada peringkat
- Kriteria 5 SwotDokumen2 halamanKriteria 5 SwotWiwit Zuriati UnoBelum ada peringkat
- Rangkuman LED Kriteria 5Dokumen1 halamanRangkuman LED Kriteria 5Wiwit Zuriati UnoBelum ada peringkat
- 5 EksipienDokumen2 halaman5 EksipienWiwit Zuriati UnoBelum ada peringkat
- Inflammatory Bowel DiseaseDokumen4 halamanInflammatory Bowel DiseaseWiwit Zuriati UnoBelum ada peringkat
- Abdulrahman Hanapi Analis MikrobiologiDokumen1 halamanAbdulrahman Hanapi Analis MikrobiologiWiwit Zuriati UnoBelum ada peringkat
- Abdulrahman Hanapi Analis MikrobiologiDokumen1 halamanAbdulrahman Hanapi Analis MikrobiologiWiwit Zuriati UnoBelum ada peringkat
- Osce 3Dokumen1 halamanOsce 3Wiwit Zuriati UnoBelum ada peringkat
- 5 EksipienDokumen2 halaman5 EksipienWiwit Zuriati UnoBelum ada peringkat
- IkoikoDokumen1 halamanIkoikoMuhammad RaisBelum ada peringkat
- Osce 3Dokumen1 halamanOsce 3Wiwit Zuriati UnoBelum ada peringkat
- Pengantar Blok Obat TradisionalDokumen35 halamanPengantar Blok Obat Tradisionalrumondang ruminingsihBelum ada peringkat
- Kontaminasi Obat Herbal Pert2Dokumen17 halamanKontaminasi Obat Herbal Pert2Wiwit Zuriati UnoBelum ada peringkat
- P1 UTS - Pengantar Obat Bahan AlamDokumen24 halamanP1 UTS - Pengantar Obat Bahan AlamDini NurchasanahBelum ada peringkat
- Analisa KosmetikDokumen15 halamanAnalisa Kosmetikrizka100% (1)
- Surat Determinasi Kementerian Pendidikan Dan KebudayaanDokumen2 halamanSurat Determinasi Kementerian Pendidikan Dan KebudayaanWiwit Zuriati UnoBelum ada peringkat
- Surat PernyataanDokumen1 halamanSurat PernyataanWiwit Zuriati UnoBelum ada peringkat
- Surat PernyataanDokumen1 halamanSurat PernyataanWiwit Zuriati UnoBelum ada peringkat
- ArtikelDokumen6 halamanArtikelWiwit Zuriati UnoBelum ada peringkat
- Lkti DMDokumen19 halamanLkti DMWiwit Zuriati UnoBelum ada peringkat