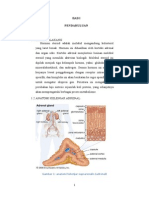
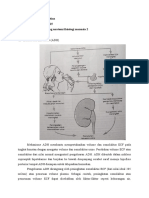
Anda mungkin juga menyukai
- AdrenalDokumen10 halamanAdrenalHimawan Dwi PrakosoBelum ada peringkat
- Makalah Cushing - Kelenjar Adrenal1Dokumen7 halamanMakalah Cushing - Kelenjar Adrenal1Qisthina Novita QuddusBelum ada peringkat
- Hormon Korteks AdrenalDokumen9 halamanHormon Korteks AdrenalCynthia Laurent SantosoBelum ada peringkat
- Hormon Kortisol NewDokumen8 halamanHormon Kortisol NewRahma Cita HalidaBelum ada peringkat
- GlikolisisDokumen14 halamanGlikolisisHensen TobingBelum ada peringkat
- Biosintesis KolesterolDokumen6 halamanBiosintesis Kolesteroldellad16Belum ada peringkat
- Kelenjar Adrenal FisiologiDokumen20 halamanKelenjar Adrenal Fisiologikeziaauresha100% (1)
- Hormon Korteks AdrenalDokumen3 halamanHormon Korteks AdrenalIsti Insani AmaliaBelum ada peringkat
- Task Reading Hormon SteroidDokumen23 halamanTask Reading Hormon SteroidAmalia S BurhanBelum ada peringkat
- Cushing SyndromeDokumen23 halamanCushing SyndromedediunggulBelum ada peringkat
- KARBOHIDRATDokumen105 halamanKARBOHIDRATelsmeeBelum ada peringkat
- Fungsi RE1Dokumen16 halamanFungsi RE1Elfa RahmadaniBelum ada peringkat
- Hormon Korteks AdrenalDokumen26 halamanHormon Korteks AdrenalIis IstikomahBelum ada peringkat
- Resume Respirasi - Kelompok 1 - Kelas JDokumen33 halamanResume Respirasi - Kelompok 1 - Kelas JI Dewa Gede Pramuja SuryanaBelum ada peringkat
- RESPIRASIDokumen23 halamanRESPIRASIMuhammad FitriansyahBelum ada peringkat
- Biokimia (Hormon Korteks Ginjal)Dokumen20 halamanBiokimia (Hormon Korteks Ginjal)abu toatBelum ada peringkat
- RespirasiDokumen24 halamanRespirasiJulia WatiBelum ada peringkat
- Korteks AdrenalDokumen34 halamanKorteks AdrenalvnBelum ada peringkat
- Hasil DK 2 Pemicu 1Dokumen28 halamanHasil DK 2 Pemicu 1Anggini Tsamaratul QolbyBelum ada peringkat
- LTM 1 BioenergetikaDokumen9 halamanLTM 1 BioenergetikarizkaBelum ada peringkat
- Respirasi SelDokumen4 halamanRespirasi SelAmelia FirdaBelum ada peringkat
- Tugas Botani FarmasiDokumen13 halamanTugas Botani FarmasisipbpokerBelum ada peringkat
- Adenosin TriPhosphateDokumen4 halamanAdenosin TriPhosphateaasatityBelum ada peringkat
- Pengertian KatabolismeDokumen11 halamanPengertian KatabolismeAnathasya SalamatBelum ada peringkat
- Biokimia Darah Dan Energi Otot JantungDokumen6 halamanBiokimia Darah Dan Energi Otot JantungTeresia Maharani100% (1)
- Kelompok 2Dokumen14 halamanKelompok 2Ria Ayu MarvinaBelum ada peringkat
- Energi Sel 2Dokumen32 halamanEnergi Sel 2Lisa AndriyaniBelum ada peringkat
- OxytocinDokumen5 halamanOxytocinDwi agustinaBelum ada peringkat
- Metabolisme Dan Respirasi SelDokumen19 halamanMetabolisme Dan Respirasi SelDewa DehBelum ada peringkat
- ATP Membawa Energi Dari Reaksi Katabolik Ke Reaksi AnabolikDokumen9 halamanATP Membawa Energi Dari Reaksi Katabolik Ke Reaksi AnabolikFahmingggowBelum ada peringkat
- Biokimia DarahDokumen8 halamanBiokimia DarahMareta AnggeliaBelum ada peringkat
- Kuis Biokimia (Metabolisme Karbohidrat) Mohammad Arif Arsaf 1900025Dokumen8 halamanKuis Biokimia (Metabolisme Karbohidrat) Mohammad Arif Arsaf 1900025Mohammad ArifBelum ada peringkat
- Biok Jantung Uho Dak18Dokumen123 halamanBiok Jantung Uho Dak18Fadel Rajab NugrahaBelum ada peringkat
- Sintesis Hormon SteroidDokumen4 halamanSintesis Hormon SteroidZulfahmi MusllimBelum ada peringkat
- Anti Diuretika HormonDokumen16 halamanAnti Diuretika HormonAsfar SyafarBelum ada peringkat
- Biokimia Nutrisi Pertemuan 2Dokumen26 halamanBiokimia Nutrisi Pertemuan 2Eka NurjannahBelum ada peringkat
- Basic Biology - St. Indah Maharani-220107512007Dokumen20 halamanBasic Biology - St. Indah Maharani-220107512007St. Indah MaharaniBelum ada peringkat
- Basic BiologyDokumen14 halamanBasic BiologySt. Indah MaharaniBelum ada peringkat
- Catatan Biologi Tugas Ke6Dokumen11 halamanCatatan Biologi Tugas Ke6Salsa AraBelum ada peringkat
- SteroidogenesisDokumen11 halamanSteroidogenesisnisa100% (3)
- Metabolisme KarbohidratDokumen15 halamanMetabolisme KarbohidratEd WingsBelum ada peringkat
- Anatomi Dan Fisiologi Kelenjar AdrenalDokumen10 halamanAnatomi Dan Fisiologi Kelenjar AdrenaldilaladyanaBelum ada peringkat
- Metabolisme Energi Otot JantungDokumen6 halamanMetabolisme Energi Otot JantungKacamata BagusBelum ada peringkat
- Reducing PowerDokumen10 halamanReducing Powerday_y464miBelum ada peringkat
- Kelenjar AdrenalDokumen19 halamanKelenjar AdrenalBts WifeBelum ada peringkat
- Sintesis ATPDokumen5 halamanSintesis ATPtantri febriana putriBelum ada peringkat
- Cardiac MetabolismeDokumen83 halamanCardiac MetabolismeJaksen ArrBelum ada peringkat
- EpinefrinDokumen4 halamanEpinefrinIMa POenya100% (1)
- LKM Biomedik DasarDokumen5 halamanLKM Biomedik DasarUlul IsmiBelum ada peringkat
- Metabolisme KarbohidratDokumen34 halamanMetabolisme KarbohidratPutri PutriBelum ada peringkat
- Metabolisme SelDokumen22 halamanMetabolisme SelNisachachaBelum ada peringkat
- Bioenergetika PresentasiDokumen42 halamanBioenergetika PresentasiYohana VeronicaBelum ada peringkat
- Asam NukleatDokumen8 halamanAsam NukleatNadia NovitaBelum ada peringkat
- RESPIRASIDokumen22 halamanRESPIRASIAndriessa PrameswaraBelum ada peringkat
- Biochemistry of Respiratory SystemDokumen153 halamanBiochemistry of Respiratory SystemNurhaya RaffyBelum ada peringkat
- Metabolisme GlukosaDokumen6 halamanMetabolisme GlukosaAbdullah RamadhanBelum ada peringkat
- BIP Case 2Dokumen2 halamanBIP Case 2TtrBelum ada peringkat
- Cover LaporanDokumen3 halamanCover LaporanTtrBelum ada peringkat
- Pertanyaan Case 4 EMSDokumen5 halamanPertanyaan Case 4 EMSTtrBelum ada peringkat
- Cover LaporanDokumen3 halamanCover LaporanTtrBelum ada peringkat
- LI Komedogenesis Dan Acne SeverityDokumen3 halamanLI Komedogenesis Dan Acne SeverityTtrBelum ada peringkat
- CoverDokumen4 halamanCoverTtrBelum ada peringkat
- PEDIGREExDokumen8 halamanPEDIGREExTtrBelum ada peringkat
- 1Dokumen7 halaman1TtrBelum ada peringkat
- Chapter IIDokumen19 halamanChapter IIPratamasari InsaniBelum ada peringkat
- 1Dokumen7 halaman1TtrBelum ada peringkat
- Arsitektur Sel Dan Struktur Membran (Dr. AB)Dokumen45 halamanArsitektur Sel Dan Struktur Membran (Dr. AB)TtrBelum ada peringkat
- Kelenjar TiroidDokumen7 halamanKelenjar TiroidTtrBelum ada peringkat
- Kelenjar TiroidDokumen7 halamanKelenjar TiroidTtrBelum ada peringkat
- 674 1255 1 SMDokumen9 halaman674 1255 1 SMristaBelum ada peringkat
- Pertanyaan Case 1 Biomedik 2Dokumen4 halamanPertanyaan Case 1 Biomedik 2TtrBelum ada peringkat
- Effective ListeningDokumen4 halamanEffective ListeningTtrBelum ada peringkat
- Sumber Ajaran IslamDokumen8 halamanSumber Ajaran IslamTtrBelum ada peringkat
- Gambar PDFDokumen4 halamanGambar PDFTtrBelum ada peringkat
- Pertanyaan Case 1 Biomedik 2Dokumen4 halamanPertanyaan Case 1 Biomedik 2TtrBelum ada peringkat