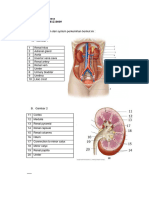
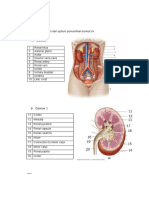
Anda mungkin juga menyukai
- Case HipokalemiaDokumen26 halamanCase HipokalemiaridhoBelum ada peringkat
- Oxytocin Dan FSHDokumen6 halamanOxytocin Dan FSHDwi agustinaBelum ada peringkat
- Perubahan Struktur SelDokumen5 halamanPerubahan Struktur SelSulchan Chris Wardana100% (2)
- Jaringan Perifer TermasuDokumen14 halamanJaringan Perifer TermasuEko AviantoBelum ada peringkat
- GabunganDokumen13 halamanGabunganDede IskandarBelum ada peringkat
- Sk1 Blok EndokrinDokumen20 halamanSk1 Blok EndokrinDimasAdriyonoWBelum ada peringkat
- HIPERALDOSTERONISMEDokumen3 halamanHIPERALDOSTERONISMESarnings SalamBelum ada peringkat
- AtikaDokumen10 halamanAtikaMegaBelum ada peringkat
- Jagat Saputra Tugas 3 BiokimiaDokumen7 halamanJagat Saputra Tugas 3 BiokimiaJagatBelum ada peringkat
- SK 3 Lo Proses Pembentukan UrinDokumen12 halamanSK 3 Lo Proses Pembentukan UrinRessita FanniaBelum ada peringkat
- PBL Blok11 Metabolik EndokrinDokumen30 halamanPBL Blok11 Metabolik EndokrinputuadiputraBelum ada peringkat
- Kelenjar Adrenal FisiologiDokumen20 halamanKelenjar Adrenal Fisiologikeziaauresha100% (1)
- Tri Essi Wahyuni - Gagal GinjalDokumen12 halamanTri Essi Wahyuni - Gagal Ginjalnurrifqoh sariBelum ada peringkat
- LP - Hipokalemia OkeDokumen26 halamanLP - Hipokalemia OkeMahfuz Azianoor50% (2)
- Biokimia Sistem KardiovaskulerDokumen10 halamanBiokimia Sistem KardiovaskulerOcda RavendraBelum ada peringkat
- Anti Diuretika HormonDokumen16 halamanAnti Diuretika HormonAsfar SyafarBelum ada peringkat
- Studi Kasus Icu DaringDokumen6 halamanStudi Kasus Icu DaringAnnisa HasnaBelum ada peringkat
- Resume Skenario 2 Tutorial HDokumen26 halamanResume Skenario 2 Tutorial HclarestaBelum ada peringkat
- Keseimbangan ElektrolitDokumen33 halamanKeseimbangan ElektrolitPUTRI LITAPRIANI100% (1)
- Korteks AdrenalDokumen34 halamanKorteks AdrenalvnBelum ada peringkat
- Referat Cushing SyndromeDokumen21 halamanReferat Cushing SyndromeTito Haposan Tobing100% (1)
- Blok 11 PBLDokumen31 halamanBlok 11 PBLHendra SusantoBelum ada peringkat
- TUGAS BMB 8 MaretDokumen3 halamanTUGAS BMB 8 MaretpipinyaquokkaBelum ada peringkat
- Review ArtikelDokumen18 halamanReview ArtikelNalendra Tri WidhianartoBelum ada peringkat
- Fisiologi HeparDokumen7 halamanFisiologi HeparmajemukaBelum ada peringkat
- Bab I HipokalemiaDokumen14 halamanBab I HipokalemiaReni April AnaBelum ada peringkat
- TGS Nusria (F201901041) Farmakologi2Dokumen6 halamanTGS Nusria (F201901041) Farmakologi2NusriaBelum ada peringkat
- Laporan Praktikum Fisiologi Kelompok B3Dokumen29 halamanLaporan Praktikum Fisiologi Kelompok B3Dewinta PutriBelum ada peringkat
- E1A021068 - Zyahwa Sabina MRP - Biokimia P3Dokumen11 halamanE1A021068 - Zyahwa Sabina MRP - Biokimia P3Zahwa SabinaBelum ada peringkat
- Anestesi SyokDokumen54 halamanAnestesi SyokwennyBelum ada peringkat
- Penugasan Blok Uropoetik1Dokumen114 halamanPenugasan Blok Uropoetik1vha_moisBelum ada peringkat
- Syok Hipovolemik Kel 3Dokumen12 halamanSyok Hipovolemik Kel 3Fitriyani WidiaBelum ada peringkat
- Biokimia Sistem KardiovaskulerDokumen11 halamanBiokimia Sistem KardiovaskulerAndi Nurul IlmiBelum ada peringkat
- Mekanisme Pembentukan UrineDokumen5 halamanMekanisme Pembentukan UrineIko Nilam SariBelum ada peringkat
- Hormon Kortisol NewDokumen8 halamanHormon Kortisol NewRahma Cita HalidaBelum ada peringkat
- Terjemahan Fitzpatrick Halaman 717 720Dokumen8 halamanTerjemahan Fitzpatrick Halaman 717 720Intan Karnina PutriBelum ada peringkat
- Laporan BiokimDokumen25 halamanLaporan Biokimdevi ainiahBelum ada peringkat
- HiperkalemiaDokumen22 halamanHiperkalemiaindra_obgBelum ada peringkat
- Fisiologi PankreasDokumen24 halamanFisiologi PankreasMawar Izzati NadillaBelum ada peringkat
- Tipus Ikan GuppyDokumen4 halamanTipus Ikan GuppyMuhammad LukmanBelum ada peringkat
- Atn-Akut Penting!!Dokumen41 halamanAtn-Akut Penting!!scheinmessakhBelum ada peringkat
- Cushing SyndromeDokumen23 halamanCushing SyndromedediunggulBelum ada peringkat
- AdrenalDokumen10 halamanAdrenalHimawan Dwi PrakosoBelum ada peringkat
- Via Lattansa (TUGAS KMB II 2A S1Kep)Dokumen6 halamanVia Lattansa (TUGAS KMB II 2A S1Kep)via lattansaBelum ada peringkat
- Anatomi Fisiologi Kelenjar AdrenalDokumen9 halamanAnatomi Fisiologi Kelenjar Adrenal-Rie Chen-100% (1)
- Fisiologi UrinDokumen41 halamanFisiologi UrinichtiyaBelum ada peringkat
- Jenis Dan Proses Second MessengerDokumen8 halamanJenis Dan Proses Second MessengerSiska NurfauziaBelum ada peringkat
- Metabolisme KarbohidratDokumen27 halamanMetabolisme KarbohidratDyah Rahayu SawitriBelum ada peringkat
- Asri Awalia Ramjani (Sistem Perkemihan Modul 6)Dokumen6 halamanAsri Awalia Ramjani (Sistem Perkemihan Modul 6)Risma Yuliana PratiwiBelum ada peringkat
- HORMONDokumen12 halamanHORMONbuti_azhaliBelum ada peringkat
- LP DM GangrenDokumen18 halamanLP DM GangrenNurul Fahmi Rizka LailyBelum ada peringkat
- Fungsi Vaskular HatiDokumen4 halamanFungsi Vaskular Hatiteguh sBelum ada peringkat
- Referat Eunice - Gagal Ginjal AkutDokumen16 halamanReferat Eunice - Gagal Ginjal AkutElisabethJaniceRusliBelum ada peringkat
- Keseimbangan Cairan Dan ElektrolitDokumen5 halamanKeseimbangan Cairan Dan ElektrolitHanny FadhilaBelum ada peringkat
- Metabolisme Karbohidrat PDFDokumen33 halamanMetabolisme Karbohidrat PDFDesy Sasana Utami PutriBelum ada peringkat
- Gangguan ElektrolitDokumen23 halamanGangguan ElektrolitasriBelum ada peringkat
- PengantarDokumen5 halamanPengantarauliya nidaBelum ada peringkat
- UNDANGDokumen5 halamanUNDANGDwi agustinaBelum ada peringkat
- Forensik 2Dokumen3 halamanForensik 2Dwi agustinaBelum ada peringkat
- Forensik 1Dokumen4 halamanForensik 1Dwi agustinaBelum ada peringkat
- Forensik 3Dokumen3 halamanForensik 3Dwi agustinaBelum ada peringkat
- Forensik 2Dokumen3 halamanForensik 2Dwi agustinaBelum ada peringkat
- Forensik 3Dokumen3 halamanForensik 3Dwi agustinaBelum ada peringkat
- Forensik 1Dokumen4 halamanForensik 1Dwi agustinaBelum ada peringkat
- Catatan Covid-19Dokumen5 halamanCatatan Covid-19Dwi agustinaBelum ada peringkat
- Heptapeptide-Loaded Solid Lipid Nanoparticles For Cosmetic Anti-Aging Applications (INd)Dokumen12 halamanHeptapeptide-Loaded Solid Lipid Nanoparticles For Cosmetic Anti-Aging Applications (INd)Dwi agustinaBelum ada peringkat
- AnginaDokumen28 halamanAnginaDwi agustinaBelum ada peringkat
- BAB-9 Enzim PPT UPIDokumen16 halamanBAB-9 Enzim PPT UPIdadan_supardan_3Belum ada peringkat
- Gangguan Nutrisi 1Dokumen6 halamanGangguan Nutrisi 1Dwi agustinaBelum ada peringkat
- Arti Covid-19Dokumen20 halamanArti Covid-19Dwi agustinaBelum ada peringkat
- Partisi Manusia Adalah Urutan Kompleks Peristiwa Yang Menyimpulkan Dalam Proses Terkoordinasi Dari Dilastasi Serviks Dan Kontraksi RahimDokumen7 halamanPartisi Manusia Adalah Urutan Kompleks Peristiwa Yang Menyimpulkan Dalam Proses Terkoordinasi Dari Dilastasi Serviks Dan Kontraksi RahimDwi agustinaBelum ada peringkat
- Buku 1Dokumen9 halamanBuku 1Dwi agustinaBelum ada peringkat
- Sistem Endokrin Dan MetabolikDokumen36 halamanSistem Endokrin Dan MetabolikDwi agustinaBelum ada peringkat
- LIPIDDokumen6 halamanLIPIDDwi agustinaBelum ada peringkat
- Resume Tetes MataDokumen2 halamanResume Tetes MataDwi agustinaBelum ada peringkat
- Bab 2 Dan Bab 3Dokumen24 halamanBab 2 Dan Bab 3Dwi agustinaBelum ada peringkat
- Golongan TerpenoidDokumen24 halamanGolongan TerpenoidDwi agustinaBelum ada peringkat
- Buku 2Dokumen3 halamanBuku 2Dwi agustinaBelum ada peringkat
- AssessmentDokumen2 halamanAssessmentDwi agustinaBelum ada peringkat
- Format Laporan Atau Jurnal Praktikum Di BiologiDokumen19 halamanFormat Laporan Atau Jurnal Praktikum Di BiologiDwi agustinaBelum ada peringkat
- Hasmat RevisiDokumen2 halamanHasmat RevisiDwi agustinaBelum ada peringkat
- AssessmentDokumen2 halamanAssessmentDwi agustinaBelum ada peringkat
- Rangkuman TentaDokumen3 halamanRangkuman TentaDwi agustinaBelum ada peringkat
- AssessmentDokumen2 halamanAssessmentDwi agustinaBelum ada peringkat
- Poin FDokumen7 halamanPoin FDwi agustinaBelum ada peringkat