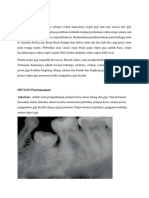
Anda mungkin juga menyukai
- PertumbuhanGigiDokumen14 halamanPertumbuhanGigiLita DamafitraBelum ada peringkat
- Atinul K. Erupsi Gigi Drg. Melv SP - KGDokumen35 halamanAtinul K. Erupsi Gigi Drg. Melv SP - KGAtinul KulsumBelum ada peringkat
- ANALISIS PROFILDokumen70 halamanANALISIS PROFILKurnia WijayantiBelum ada peringkat
- Laporan Tutorial Skenario 2Dokumen30 halamanLaporan Tutorial Skenario 2Afif SuryaBelum ada peringkat
- SK 4 TentirDokumen63 halamanSK 4 TentirAnonymous d5LNpKzBelum ada peringkat
- Kelainan Petumbuhan Dan Perkembangan Dentofasial 2Dokumen89 halamanKelainan Petumbuhan Dan Perkembangan Dentofasial 2Kevin Marselinus100% (1)
- Skenario 2 Hums FixDokumen18 halamanSkenario 2 Hums FixSindystyaBelum ada peringkat
- Struktur Gigi SulungDokumen35 halamanStruktur Gigi SulungFarda MadinBelum ada peringkat
- Pengunyahan Danb PenelananDokumen12 halamanPengunyahan Danb PenelananWulan Oktaviani0% (1)
- Kelompok 2 FasialDokumen6 halamanKelompok 2 FasialTimothy MedinaBelum ada peringkat
- Gerakan TMJDokumen5 halamanGerakan TMJJasmine R D MBelum ada peringkat
- Mekanisme Dentinogenesis Dan Penyembuhan PulpaDokumen2 halamanMekanisme Dentinogenesis Dan Penyembuhan PulpaBenita AryaniBelum ada peringkat
- Petunjuk Praktikum Fungsi Sistem Stomatognasi - 1Dokumen8 halamanPetunjuk Praktikum Fungsi Sistem Stomatognasi - 1EndungBelum ada peringkat
- Petunjuk Praktikum 01: Blok Fungsi Sistem StomatognatiDokumen11 halamanPetunjuk Praktikum 01: Blok Fungsi Sistem StomatognatiReza PahleviBelum ada peringkat
- Biomekanika TMJDokumen1 halamanBiomekanika TMJIntan Kamala AisyiahBelum ada peringkat
- Laporan Tutorial Skenario 4Dokumen30 halamanLaporan Tutorial Skenario 4Maria DevithaBelum ada peringkat
- Morfologi Gigi Desidui Dan Gigi Permanen - Final - Normal - Bab 1Dokumen9 halamanMorfologi Gigi Desidui Dan Gigi Permanen - Final - Normal - Bab 1DIen ARchBelum ada peringkat
- AmelogenesisDokumen5 halamanAmelogenesisFachriza AfriBelum ada peringkat
- Laporan Skenario 2Dokumen22 halamanLaporan Skenario 2Dhesyarmani P. RothschildiBelum ada peringkat
- 3 Tumbuh Kembang Cavum Nasi Dan Cavum OrisDokumen12 halaman3 Tumbuh Kembang Cavum Nasi Dan Cavum OrisKhairunnisa RasyidinBelum ada peringkat
- Perkembangan Oklusi Dan Relasi RahangDokumen3 halamanPerkembangan Oklusi Dan Relasi RahangParamita Ekadeva Sari DBelum ada peringkat
- Tutorial Modul 2 Blok 5Dokumen19 halamanTutorial Modul 2 Blok 5anisa raudhatul husnaBelum ada peringkat
- Anatomi PulpaDokumen24 halamanAnatomi PulpachelsyBelum ada peringkat
- Kelompok 1Dokumen34 halamanKelompok 1Anonymous WHFyFWBelum ada peringkat
- Materi Modul 4 Blok 7 WordDokumen3 halamanMateri Modul 4 Blok 7 WordArfrindaBelum ada peringkat
- Motekar Blok 2 DeyuDokumen18 halamanMotekar Blok 2 DeyuIkbarBelum ada peringkat
- Blok 3 Skenario 1Dokumen19 halamanBlok 3 Skenario 1yufidaBelum ada peringkat
- VARIASI ERUKSI GIGIDokumen3 halamanVARIASI ERUKSI GIGIBima AjiBelum ada peringkat
- Dimensi Vertikal GTDokumen11 halamanDimensi Vertikal GTdewi zulianaBelum ada peringkat
- Pengaruh Nutrisi Terhadap Jaringan Lunak Mulut Fix !Dokumen17 halamanPengaruh Nutrisi Terhadap Jaringan Lunak Mulut Fix !Sri Rahmawati0% (3)
- Anatomy Gigi - FKG 2010Dokumen82 halamanAnatomy Gigi - FKG 2010Dedi MaryadiBelum ada peringkat
- Tahap Pertumbuhan GigiDokumen2 halamanTahap Pertumbuhan GigiLiaBelum ada peringkat
- Perkembangan Kelenjar SalivaDokumen3 halamanPerkembangan Kelenjar SalivaMarina RosyanaBelum ada peringkat
- Pertumbuhan GigiDokumen21 halamanPertumbuhan GigiAniska Cattleya Shara100% (2)
- SERABUT PERIODONTALDokumen22 halamanSERABUT PERIODONTALArmansyah PutraBelum ada peringkat
- Erupsi GigiDokumen10 halamanErupsi GigissssBelum ada peringkat
- Gigi DewasaDokumen45 halamanGigi DewasaClaraBelum ada peringkat
- Anatomi Dan Histologi Temporomandibular JointDokumen5 halamanAnatomi Dan Histologi Temporomandibular JointbagasBelum ada peringkat
- LAPORAN PRAKTIKUM BIOLOGI ORAL p5 Blok 6 - b3Dokumen30 halamanLAPORAN PRAKTIKUM BIOLOGI ORAL p5 Blok 6 - b3dafaBelum ada peringkat
- TinggiBadan20TahunDokumen2 halamanTinggiBadan20TahunNengah Vindi100% (1)
- Laporan Modul 4Dokumen15 halamanLaporan Modul 4kiidouble100% (2)
- PertumbuhanWajahDokumen14 halamanPertumbuhanWajahKevin Clara DeaBelum ada peringkat
- PERKEMBANGAN GIGIDokumen4 halamanPERKEMBANGAN GIGIDewiayu Shs DewangBelum ada peringkat
- StogmatonatikDokumen16 halamanStogmatonatikZahra Najmi Afifah100% (1)
- TMJDokumen13 halamanTMJPutri Silvia100% (1)
- Praktikum Kegawatdaruratan Medic DentalDokumen18 halamanPraktikum Kegawatdaruratan Medic Dentaladlyfaris100% (1)
- PERTUMBUHANDokumen38 halamanPERTUMBUHANRizky PutriBelum ada peringkat
- Kelainan GigiDokumen11 halamanKelainan GigiSupri BriyantiniBelum ada peringkat
- Proses Pembentukan Dentin, Kontrol, Isolasi - EvaDokumen52 halamanProses Pembentukan Dentin, Kontrol, Isolasi - EvaEva DewiBelum ada peringkat
- FaktorGenetikLingkunganDokumen21 halamanFaktorGenetikLingkunganbellaBelum ada peringkat
- ErupsiGigiDokumen6 halamanErupsiGigimuga restunaesha100% (1)
- Pertumbuhan Dan Perkembangan Wajah PostnatalDokumen14 halamanPertumbuhan Dan Perkembangan Wajah Postnatalanisa raudhatul husnaBelum ada peringkat
- Pengembangan Kelenjar SalivaDokumen43 halamanPengembangan Kelenjar SalivaSariBelum ada peringkat
- Pemicu-1 Meili Kel 12 SHDokumen167 halamanPemicu-1 Meili Kel 12 SHwennyBelum ada peringkat
- Pembentukan BlastokistaDokumen2 halamanPembentukan BlastokistaNaufal100% (1)
- Embryology and Development (Article) Author G Kurkchubasche Id PDFDokumen6 halamanEmbryology and Development (Article) Author G Kurkchubasche Id PDFdava fathiyaBelum ada peringkat
- GASTRULASIDokumen3 halamanGASTRULASIana saniaBelum ada peringkat
- PERKEMBANGAN ORGAN EMBRIO KATAKDokumen6 halamanPERKEMBANGAN ORGAN EMBRIO KATAKUmmi AmaliaBelum ada peringkat
- Biologi Reproduksi Pada HewanDokumen20 halamanBiologi Reproduksi Pada Hewannila_sdBelum ada peringkat
- S. Integumentum-Kuliah (DR - Arif)Dokumen55 halamanS. Integumentum-Kuliah (DR - Arif)Andri Adma WijayaBelum ada peringkat
- BAB 2 ZincDokumen3 halamanBAB 2 ZincMuhammad Harmen Reza SiregarBelum ada peringkat
- ISIDokumen17 halamanISIMuhammad Harmen Reza SiregarBelum ada peringkat
- ZincDokumen1 halamanZincMuhammad Harmen Reza SiregarBelum ada peringkat
- Paper OsteosarkomaDokumen20 halamanPaper OsteosarkomaMuhammad Harmen Reza SiregarBelum ada peringkat
- Bab 1Dokumen3 halamanBab 1Muhammad Harmen Reza SiregarBelum ada peringkat
- HiperprolaktinemiaDokumen18 halamanHiperprolaktinemiaMuhammad Harmen Reza SiregarBelum ada peringkat
- Jadwal Jaga JuniDokumen1 halamanJadwal Jaga JuniMuhammad Harmen Reza SiregarBelum ada peringkat
- Bab 1Dokumen3 halamanBab 1Muhammad Harmen Reza SiregarBelum ada peringkat
- AntiaritmiaDokumen21 halamanAntiaritmiaMuhammad Harmen Reza SiregarBelum ada peringkat
- 11 Pengujian Hipotesis Rata Rata Dua PopulasiDokumen8 halaman11 Pengujian Hipotesis Rata Rata Dua PopulasiDikdik Pramiady PutraBelum ada peringkat
- Daftar IsiDokumen34 halamanDaftar IsiMuhammad Harmen Reza SiregarBelum ada peringkat
- LAPKAS Anestesi Intususepsi, InvaginasiDokumen44 halamanLAPKAS Anestesi Intususepsi, InvaginasiMuhammad Harmen Reza SiregarBelum ada peringkat
- ZincDokumen1 halamanZincMuhammad Harmen Reza SiregarBelum ada peringkat
- Journal Reading CoverDokumen10 halamanJournal Reading CoverMuhammad Harmen Reza SiregarBelum ada peringkat
- Fisiologi Kardiovaskuler Dan AnestesiDokumen53 halamanFisiologi Kardiovaskuler Dan AnestesiD'ian See TreeDhiBelum ada peringkat
- TraumaKepalaDokumen33 halamanTraumaKepalaMuhammad Harmen Reza SiregarBelum ada peringkat
- TBR Pak DosDokumen19 halamanTBR Pak DosMuhammad Harmen Reza SiregarBelum ada peringkat
- TBR Pak DosDokumen23 halamanTBR Pak DosMuhammad Harmen Reza SiregarBelum ada peringkat
- Hipertensi Pada KehamilanDokumen24 halamanHipertensi Pada KehamilanMuhammad Harmen Reza SiregarBelum ada peringkat
- Daftar IsiDokumen2 halamanDaftar IsiMuhammad Harmen Reza SiregarBelum ada peringkat
- Chapter IDokumen3 halamanChapter IMuhammad Harmen Reza SiregarBelum ada peringkat
- TBR Pak DosDokumen19 halamanTBR Pak DosMuhammad Harmen Reza SiregarBelum ada peringkat
- Hipertensi Pada KehamilanDokumen24 halamanHipertensi Pada KehamilanMuhammad Harmen Reza SiregarBelum ada peringkat
- Chapter IIDokumen22 halamanChapter IIMuhammad Harmen Reza SiregarBelum ada peringkat
- Lapkas JadiDokumen41 halamanLapkas JadiMuhammad Harmen Reza SiregarBelum ada peringkat
- BoiiiDokumen11 halamanBoiiiMuhammad Harmen Reza SiregarBelum ada peringkat
- Anatomi Kelenjar TiroidDokumen16 halamanAnatomi Kelenjar TiroidMuhammad Harmen Reza SiregarBelum ada peringkat
- ReferenceDokumen3 halamanReferenceMuhammad Harmen Reza SiregarBelum ada peringkat
- Nomor ICDDokumen19 halamanNomor ICDMuhammad Harmen Reza SiregarBelum ada peringkat