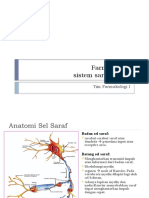
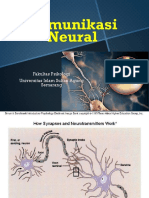
Anda mungkin juga menyukai
- Penghantar Impuls Didalam Tubuh Dan Transmisi SinapsisDokumen5 halamanPenghantar Impuls Didalam Tubuh Dan Transmisi SinapsisAshri Margiana89% (9)
- Makalah Penghantar Impuls Dalam TubuhDokumen11 halamanMakalah Penghantar Impuls Dalam TubuhDa Suprem100% (4)
- Penghantar Impuls Didalam Tubuh Dan Transmisi SinapsisDokumen31 halamanPenghantar Impuls Didalam Tubuh Dan Transmisi SinapsisCeni100% (3)
- SINAPSISDokumen29 halamanSINAPSISdevina putri100% (1)
- NeurotransmitterDokumen2 halamanNeurotransmitterJustMahaSiswaBelum ada peringkat
- Potensial Aksi Dan SynapsDokumen9 halamanPotensial Aksi Dan SynapsOgi NurulBelum ada peringkat
- Bahan SarafDokumen39 halamanBahan SarafFabianus TegarBelum ada peringkat
- Penghantaran Impuls Didalam TubuhDokumen9 halamanPenghantaran Impuls Didalam TubuhLinaBelum ada peringkat
- TRANSISISINYALDokumen10 halamanTRANSISISINYALBegum TiffanyBelum ada peringkat
- Dhea dk2 Pemicu 3Dokumen16 halamanDhea dk2 Pemicu 3Ahmad NazharBelum ada peringkat
- Fatma N. Utina - Sistem SarafDokumen7 halamanFatma N. Utina - Sistem Saraffatmautina56Belum ada peringkat
- Mekanisme Impuls SarafDokumen6 halamanMekanisme Impuls SarafHendrika M UBelum ada peringkat
- Bio 9Dokumen3 halamanBio 9MayaBelum ada peringkat
- SynapsisDokumen4 halamanSynapsisadinda nisaBelum ada peringkat
- Impuls SarafDokumen8 halamanImpuls SarafVhiny ChaBelum ada peringkat
- NEUROTRANSMISI DAN KONDUKSIDokumen28 halamanNEUROTRANSMISI DAN KONDUKSItpat ciaBelum ada peringkat
- MEKANISME TRANSMISI SINAPSISDokumen5 halamanMEKANISME TRANSMISI SINAPSISKhofifah WulannorBelum ada peringkat
- KOMUNIKASI SISTEM SARAFDokumen3 halamanKOMUNIKASI SISTEM SARAFIvan Dwi KBelum ada peringkat
- NeuroDokumen27 halamanNeuroDanita Suci LestariBelum ada peringkat
- PBL Blok 6Dokumen21 halamanPBL Blok 6Piter PicalBelum ada peringkat
- Laporan Praktikum FisiologiDokumen16 halamanLaporan Praktikum FisiologiAnna AlthafunnisaBelum ada peringkat
- SINAPSISDokumen15 halamanSINAPSISDebora EvaniaBelum ada peringkat
- Sinaps Dan Integrasi NeuronDokumen3 halamanSinaps Dan Integrasi NeuronOliez D'respect100% (1)
- Makalah IBD Kelompok 3Dokumen11 halamanMakalah IBD Kelompok 3Annisa SyahabBelum ada peringkat
- The SynapsesDokumen3 halamanThe SynapsesNurul HayatBelum ada peringkat
- Esai Dr. TunjungDokumen11 halamanEsai Dr. TunjungWahyu KurniawanBelum ada peringkat
- SINAPSDokumen14 halamanSINAPSDanie AurelliaBelum ada peringkat
- Bahan SarafDokumen39 halamanBahan SarafErlan StrangersBelum ada peringkat
- Depresi dan BipolarDokumen13 halamanDepresi dan Bipolarrahayu dewiBelum ada peringkat
- 6,7,9,10Dokumen22 halaman6,7,9,10Pratiwi EkaputriBelum ada peringkat
- NEUROFISIOLOGIDokumen10 halamanNEUROFISIOLOGIEndang MunawatyBelum ada peringkat
- Komunikasi Antar Sel TerbaruDokumen10 halamanKomunikasi Antar Sel TerbaruDony ArdiantoBelum ada peringkat
- BIOLISTRIK OPTIMALDokumen15 halamanBIOLISTRIK OPTIMALSalwa RendratamaBelum ada peringkat
- Tabitha Puspaning Asmara (8881190021) - Pre-LectureDokumen5 halamanTabitha Puspaning Asmara (8881190021) - Pre-LectureTabitha Puspaning AsmaraBelum ada peringkat
- Pengantar SSP Dan SSPDokumen27 halamanPengantar SSP Dan SSPRaisa FebriyaBelum ada peringkat
- Sinaps dan neurotransmiterDokumen38 halamanSinaps dan neurotransmiterhayrachzaBelum ada peringkat
- BIOELEKTRIKDokumen33 halamanBIOELEKTRIKFika SuhartinaBelum ada peringkat
- Jantung BerdebarDokumen21 halamanJantung BerdebarWelly SuryaBelum ada peringkat
- Referat NeuroDokumen14 halamanReferat NeuroFuad HadiBelum ada peringkat
- IBD B-11 - Topik 1 - Sistem Saraf, Pengindraan, Dan MuskuloskeletalDokumen108 halamanIBD B-11 - Topik 1 - Sistem Saraf, Pengindraan, Dan Muskuloskeletalkimny parkBelum ada peringkat
- Prinsip Penghantaran ImpulsDokumen3 halamanPrinsip Penghantaran ImpulsOlivia RachelBelum ada peringkat
- Penghantaran Impuls Di Dalam TubuhDokumen21 halamanPenghantaran Impuls Di Dalam TubuhApriyani laila MaddatuBelum ada peringkat
- Jelaskan Apa Yang Dimaksud Dengan Sinaps Dan Bagimanakah Mekanisme Kerja Sinaps KimiaDokumen1 halamanJelaskan Apa Yang Dimaksud Dengan Sinaps Dan Bagimanakah Mekanisme Kerja Sinaps KimiaRizqi Layli KhusufiBelum ada peringkat
- Mekanisme Jalannya Impuls SyarafDokumen4 halamanMekanisme Jalannya Impuls SyarafAmelia RosyidaBelum ada peringkat
- Proses PA, sinaps, saraf simpatis dan parasimpatis, refleks kompleks, kaitan saraf dan hormon pituitariDokumen6 halamanProses PA, sinaps, saraf simpatis dan parasimpatis, refleks kompleks, kaitan saraf dan hormon pituitariTawang Handayani0% (1)
- SinapsisDokumen31 halamanSinapsisnadia afina ZulfaBelum ada peringkat
- Resume MateriDokumen9 halamanResume MateriAhmad JunaidiBelum ada peringkat
- Fungsi SinapsDokumen4 halamanFungsi SinapsNazlia AnjaniBelum ada peringkat
- NEUROTRANSMISIDokumen5 halamanNEUROTRANSMISIDyahAyuRisqiNilamsariBelum ada peringkat
- Farmakologi Sistem Saraf PusatDokumen98 halamanFarmakologi Sistem Saraf PusatPuri Ratna KartiniBelum ada peringkat
- Komunikasi NeuralDokumen48 halamanKomunikasi NeuralArkan NiawanBelum ada peringkat
- Akson Digolongkan Ke Dalam Tiga Kelompok Sesuai Dengan Hubungan Antara MielinasiDokumen4 halamanAkson Digolongkan Ke Dalam Tiga Kelompok Sesuai Dengan Hubungan Antara MielinasiprincessscantikBelum ada peringkat
- Transmisi Impuls SarafDokumen60 halamanTransmisi Impuls Sarafkurniaadhi100% (4)
- BiologiDokumen8 halamanBiologidesiBelum ada peringkat
- Anatomi NMJDokumen3 halamanAnatomi NMJmayo bqBelum ada peringkat
- INTEGRASI SINAP DAN NEURONDokumen10 halamanINTEGRASI SINAP DAN NEURONKristi SusantiBelum ada peringkat
- Fisiologi NeuroDokumen29 halamanFisiologi NeuroFathiyyah HaraBelum ada peringkat
- DT 1.3.1 No 4Dokumen5 halamanDT 1.3.1 No 4AnnisaBelum ada peringkat
- SOAL FISIOLOGI HEWAN DAN MANUSIADokumen9 halamanSOAL FISIOLOGI HEWAN DAN MANUSIAnandasalmaBelum ada peringkat