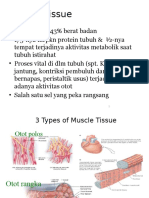
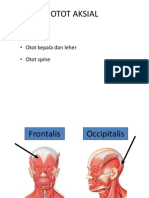
Anda mungkin juga menyukai
- Muscle Tissue-2Dokumen89 halamanMuscle Tissue-2Riandi Candra PrayogaBelum ada peringkat
- 4 - OtotDokumen31 halaman4 - OtotHabib Isima100% (1)
- Otot RangkaDokumen25 halamanOtot RangkaIlmi Octaviani GeopanyBelum ada peringkat
- Fisiologi Sistem Reproduksi 2021Dokumen61 halamanFisiologi Sistem Reproduksi 2021andra pramudiaBelum ada peringkat
- Anfis Sistem SarafDokumen28 halamanAnfis Sistem SarafMiss AisyahBelum ada peringkat
- UROGENITALDokumen40 halamanUROGENITALirmayaniBelum ada peringkat
- Materi 5 Fisiologi Nutris (NGT) LutfiDokumen46 halamanMateri 5 Fisiologi Nutris (NGT) LutfiMoh. lutfiBelum ada peringkat
- Sistem Reproduksi Manusia1Dokumen72 halamanSistem Reproduksi Manusia1Fathya Nurul AisyaBelum ada peringkat
- SISTEM PENCERNAAN MAKANAN k13Dokumen30 halamanSISTEM PENCERNAAN MAKANAN k13Davina UtamiBelum ada peringkat
- Reproduksi Pria BRDokumen27 halamanReproduksi Pria BRrisma febrianiBelum ada peringkat
- Sistem Rangka - AnfismanDokumen49 halamanSistem Rangka - AnfismanDiah Ayu PradnyaniBelum ada peringkat
- Neurofisiologi EditDokumen120 halamanNeurofisiologi EditDwiYukaBelum ada peringkat
- Persepsi Sensori PK JumDokumen60 halamanPersepsi Sensori PK JumAshley NashBelum ada peringkat
- Kuliah Xiii-Xiv - Anatomi Dan Fisiologi Sistem Panca InderaDokumen102 halamanKuliah Xiii-Xiv - Anatomi Dan Fisiologi Sistem Panca InderaIqbal Pratama NurdiansyahBelum ada peringkat
- Fisiologi Otot 2020Dokumen27 halamanFisiologi Otot 2020Nicz NazzBelum ada peringkat
- NIDASIDokumen40 halamanNIDASISiwi TrimulyaniBelum ada peringkat
- Sistem RespirasiDokumen52 halamanSistem RespirasiAnonymous NBZpegWhtBelum ada peringkat
- Sistem GerakDokumen93 halamanSistem GeraklutfimeigaBelum ada peringkat
- NeurotransmitterDokumen25 halamanNeurotransmitterAliMa'rufBelum ada peringkat
- RPS Mikrobiologi-Ung-2016Dokumen7 halamanRPS Mikrobiologi-Ung-2016Ambarwaty BilondatuBelum ada peringkat
- Sistem Persepsi Sensori: Meta Nurbaiti, S.Kep.,NsDokumen37 halamanSistem Persepsi Sensori: Meta Nurbaiti, S.Kep.,NsAnonymous fIWWmbYzrnBelum ada peringkat
- Fisiologi Otot & Mekanisme Kontraksi Otot RangkaDokumen38 halamanFisiologi Otot & Mekanisme Kontraksi Otot RangkaNarjus SafaahBelum ada peringkat
- Sistem SarafDokumen38 halamanSistem SarafKKI 2017Belum ada peringkat
- Kontraksi Dan Relaksasi OtotDokumen7 halamanKontraksi Dan Relaksasi OtotSiti FathiyaBelum ada peringkat
- Anatomi SendiDokumen30 halamanAnatomi SendiChisilia RashidBelum ada peringkat
- Sistem Saraf Dan Hormon - ShilaDokumen11 halamanSistem Saraf Dan Hormon - ShilaTjhieahya SyahvillaBelum ada peringkat
- HomeostasisDokumen31 halamanHomeostasisrafi_ptkBelum ada peringkat
- Otot AksialDokumen15 halamanOtot AksialMichelle AllisonBelum ada peringkat
- Terminologi DoneDokumen16 halamanTerminologi DoneArum Maharani LantagiBelum ada peringkat
- Istirahat Tidur PDFDokumen19 halamanIstirahat Tidur PDFRanti Ayu Kartika100% (1)
- Buku Ajar Ilmu Keperawatan DasarDokumen6 halamanBuku Ajar Ilmu Keperawatan DasarMirsya AnggrianaBelum ada peringkat
- Sistem Pencernaan MakananDokumen54 halamanSistem Pencernaan MakananFariza A GunindaBelum ada peringkat
- Hormon InsulinDokumen42 halamanHormon InsulinrealxomayBelum ada peringkat
- Makalah Resensi Psikologi KomunikasiDokumen11 halamanMakalah Resensi Psikologi KomunikasiAnisa PratiwiBelum ada peringkat
- Pengantar AnatomiDokumen60 halamanPengantar Anatominovia ayuBelum ada peringkat
- Sistem OtotDokumen32 halamanSistem OtotVennesia100% (4)
- Hipersensitivitas 11 Nov 2019 PDFDokumen48 halamanHipersensitivitas 11 Nov 2019 PDFfikowowBelum ada peringkat
- Konsep Berfikir KritisDokumen58 halamanKonsep Berfikir KritisMei SanjiwaniBelum ada peringkat
- Algoritma HipotensiDokumen1 halamanAlgoritma HipotensilenaBelum ada peringkat
- Sistem Imun Spesifik (Adaptif)Dokumen53 halamanSistem Imun Spesifik (Adaptif)Lin CrBelum ada peringkat
- Bab 3Dokumen3 halamanBab 3Zanty Rakhmania PutriBelum ada peringkat
- Dasar-Dasar AnatomiDokumen56 halamanDasar-Dasar AnatomiNuri FebrianiBelum ada peringkat
- Anatomi Fisiologi UrologiDokumen25 halamanAnatomi Fisiologi Urologiandara sudirthaBelum ada peringkat
- Fisiologi OtotDokumen35 halamanFisiologi OtotNur IrfanBelum ada peringkat
- Nutrisi Bedah - MakalahDokumen58 halamanNutrisi Bedah - MakalahAriaPratamaBelum ada peringkat
- Interaksi Antigen AntibodiDokumen7 halamanInteraksi Antigen Antibodipristy okta nilaBelum ada peringkat
- Metabolisme MikroorganismeDokumen20 halamanMetabolisme MikroorganismeRANGGA ADITYA PRADANA -Belum ada peringkat
- Fisiologi ReproduksiDokumen39 halamanFisiologi ReproduksiEva Titaniia MaulidaBelum ada peringkat
- Anatomi & Fisiologi Sistem SarafDokumen47 halamanAnatomi & Fisiologi Sistem SarafSukmawati 26Belum ada peringkat
- PETIDINDokumen3 halamanPETIDINmei indahBelum ada peringkat
- Laporan Tutorial Skenario A Blok 3Dokumen12 halamanLaporan Tutorial Skenario A Blok 3Ivan-ty FamBelum ada peringkat
- 2.4.3.6b Gastropati NSAIDDokumen22 halaman2.4.3.6b Gastropati NSAIDPutri WulandariBelum ada peringkat
- Pola Hubungan Kerja PerawatDokumen25 halamanPola Hubungan Kerja PerawatEni TrismiatiBelum ada peringkat
- Gizi Bidan 1Dokumen16 halamanGizi Bidan 1Hania PriciyliaBelum ada peringkat
- Mekanisme Kontraksi OtotDokumen6 halamanMekanisme Kontraksi OtotKamila JasmineBelum ada peringkat
- Fisiologi SelDokumen25 halamanFisiologi SelMuhammad Budi SyahputraBelum ada peringkat
- MyologiDokumen86 halamanMyologiagus safarmingBelum ada peringkat
- Fisiologi Otot 2022Dokumen28 halamanFisiologi Otot 2022Feni meliyani SwanBelum ada peringkat
- Jaringan OtotDokumen63 halamanJaringan OtotArief Nurul KurniawanBelum ada peringkat
- 7 Sistem MuskuloskeletalDokumen112 halaman7 Sistem MuskuloskeletalmithaBelum ada peringkat
- 3 Metpen-KualitatifDokumen27 halaman3 Metpen-KualitatifIka ChasaBelum ada peringkat
- Laporan Status Klinis-Trisandi KurniaDokumen32 halamanLaporan Status Klinis-Trisandi KurniaHeru PurnomoBelum ada peringkat
- Heru Purnomo-GeriatriDokumen9 halamanHeru Purnomo-GeriatriHeru PurnomoBelum ada peringkat
- Laporan Status Klinis-Trisandi KurniaDokumen32 halamanLaporan Status Klinis-Trisandi KurniaHeru PurnomoBelum ada peringkat
- Anatomi Dan Fisiologi KulitDokumen48 halamanAnatomi Dan Fisiologi KulitAnonymous w3vYureBelum ada peringkat
- Diplomasi RasulullahDokumen1 halamanDiplomasi RasulullahHeru PurnomoBelum ada peringkat
- ALZHEIMERDokumen11 halamanALZHEIMERHeru PurnomoBelum ada peringkat
- ContohDokumen29 halamanContohHeru PurnomoBelum ada peringkat
- Learning Activity - Algortima Berfikir Karya Tulis IlmiahDokumen4 halamanLearning Activity - Algortima Berfikir Karya Tulis IlmiahHeru PurnomoBelum ada peringkat