Anda mungkin juga menyukai
- Laporan Praktikum Isolasi PBMCDokumen17 halamanLaporan Praktikum Isolasi PBMCYulinar Risky KaramanBelum ada peringkat
- Pemeliharaan Biakan MurniDokumen3 halamanPemeliharaan Biakan Murniahmad kabirul rifaiBelum ada peringkat
- Aziz MIKROLINGDokumen7 halamanAziz MIKROLINGAzi Muhamad Syahreza AzizarBelum ada peringkat
- Desakan DarahDokumen15 halamanDesakan Darahimanuel kuneBelum ada peringkat
- LAPORAN Induksi Tunas Kalus (BAB 1 & BAB 2)Dokumen20 halamanLAPORAN Induksi Tunas Kalus (BAB 1 & BAB 2)Hayyu ArumsariBelum ada peringkat
- Proyek Mini Fisiologi HewanDokumen2 halamanProyek Mini Fisiologi HewanrindayuputriBelum ada peringkat
- Laporan Determinasi Drosophila - GenetikaDokumen15 halamanLaporan Determinasi Drosophila - GenetikaGhina MutiaraBelum ada peringkat
- Tinjauan Pustaka RNADokumen4 halamanTinjauan Pustaka RNAIkhwanMaulanaBelum ada peringkat
- Metode Sequencing Human GenomeDokumen23 halamanMetode Sequencing Human GenomeNani Rusyad RahmanBelum ada peringkat
- 3414 - RAPD (Random Amplified Polymorphic DNA) Kelompok 4Dokumen14 halaman3414 - RAPD (Random Amplified Polymorphic DNA) Kelompok 4elvinaBelum ada peringkat
- Makalah Transformasi SelDokumen40 halamanMakalah Transformasi Selanesco tuasuun0% (1)
- Medium Pertumbuhan MikroorganismeDokumen19 halamanMedium Pertumbuhan MikroorganismeJeremy WeaverBelum ada peringkat
- BAB II Materi Genetik-OkDokumen37 halamanBAB II Materi Genetik-OkRiska Widya LavegaBelum ada peringkat
- Diagnosa Laboratorium Paramecium SPDokumen7 halamanDiagnosa Laboratorium Paramecium SPJuchi Chaula Liza Chairani0% (1)
- Uji Biokimia BakteriDokumen15 halamanUji Biokimia Bakteriumye_558520219100% (2)
- Eubacteria 1Dokumen56 halamanEubacteria 1Wanabud abudBelum ada peringkat
- Mikro 4Dokumen12 halamanMikro 4Aji SandianaBelum ada peringkat
- RFLP CYP1A2Dokumen7 halamanRFLP CYP1A2Edo EdoBelum ada peringkat
- Pembuatan preparat irisan metode parafinDokumen18 halamanPembuatan preparat irisan metode parafinsitinBelum ada peringkat
- Mikroteknik Parafin EmbeddingDokumen14 halamanMikroteknik Parafin Embeddingcitra ayuzhaaBelum ada peringkat
- IsolasiDokumen8 halamanIsolasiroesma narulitaBelum ada peringkat
- Makalah BiomokulerDokumen28 halamanMakalah BiomokulerDeaFatikaNurhayatiBelum ada peringkat
- Laporan Praktikum Fisiologi Tumbuhan AngDokumen13 halamanLaporan Praktikum Fisiologi Tumbuhan AngGawean CoyBelum ada peringkat
- Isolasi DNA Buah Menggunakan Detergen BerbedaDokumen10 halamanIsolasi DNA Buah Menggunakan Detergen BerbedaALIEF SELLABelum ada peringkat
- SIDA_RHOMBIFOLIADokumen5 halamanSIDA_RHOMBIFOLIAMelzy Putri SaniBelum ada peringkat
- Bakteri Indikator Sanitasi Dan Keamanan Air MinumDokumen11 halamanBakteri Indikator Sanitasi Dan Keamanan Air MinumNurrahimah GafurBelum ada peringkat
- Real-Time PCRDokumen4 halamanReal-Time PCRNadea Grescia Abelia NadeaBelum ada peringkat
- Diagnosis Laboratorium Pada Penyakit InfeksiDokumen43 halamanDiagnosis Laboratorium Pada Penyakit InfeksinugrahaaryaBelum ada peringkat
- Prinsip Dan Pemanfaatan Rekayasa Genetik Dalam Produksi ObatDokumen40 halamanPrinsip Dan Pemanfaatan Rekayasa Genetik Dalam Produksi ObatFee ChocoBelum ada peringkat
- Mitek 3 SupravitalDokumen13 halamanMitek 3 SupravitalFitri suci KarlinaBelum ada peringkat
- Biologi Sel Molekuler (Teknik Isolasi Dna)Dokumen15 halamanBiologi Sel Molekuler (Teknik Isolasi Dna)Fg GAMINGBelum ada peringkat
- KULTUR PRIMERDokumen40 halamanKULTUR PRIMERRunoffBelum ada peringkat
- UJI SerologiDokumen8 halamanUJI SerologibjatmikoooBelum ada peringkat
- Dasar Teori SporaDokumen3 halamanDasar Teori SporaElsa NoviaBelum ada peringkat
- Dasar Teori PCRDokumen8 halamanDasar Teori PCRHollowfict100% (1)
- Bab II Mutasi Dan Aberasi KromosomDokumen22 halamanBab II Mutasi Dan Aberasi KromosomwennyBelum ada peringkat
- STOMATADokumen11 halamanSTOMATAPutri maulidia FitriaBelum ada peringkat
- Sintesis Protein Adalah Proses Di Mana Rantai Polipeptida Dibentuk Dari Kombinasi Kode Asam Amino Tunggal Di Dalam SelDokumen6 halamanSintesis Protein Adalah Proses Di Mana Rantai Polipeptida Dibentuk Dari Kombinasi Kode Asam Amino Tunggal Di Dalam SelWildanul HasanahBelum ada peringkat
- Pembahasan SDS PAGEDokumen7 halamanPembahasan SDS PAGEFifi FitriawatiBelum ada peringkat
- 2a Isolasi Dna RnaDokumen21 halaman2a Isolasi Dna RnayulialisnaBelum ada peringkat
- Bab MutasiDokumen3 halamanBab MutasiliviaBelum ada peringkat
- Alat Laboratorium Biologi MolekulerDokumen50 halamanAlat Laboratorium Biologi MolekulerSalwa NurafifahBelum ada peringkat
- Kode GenetikDokumen6 halamanKode GenetiknurilmaghfirohBelum ada peringkat
- Mikrobiologi - Pewarnaan EndosporaDokumen10 halamanMikrobiologi - Pewarnaan EndosporaAizIce100% (1)
- GENETIKADokumen12 halamanGENETIKAimroatun nafi'ahBelum ada peringkat
- TERMOREGULASIDokumen9 halamanTERMOREGULASISulis PujiyantiBelum ada peringkat
- Mitosis Akar BawangDokumen14 halamanMitosis Akar BawangArif Rachmatullah100% (1)
- Praktikum Suhu Tubuh BM 2Dokumen4 halamanPraktikum Suhu Tubuh BM 2tanBelum ada peringkat
- Biosintesis Karbohidrat - RisyadDokumen29 halamanBiosintesis Karbohidrat - RisyadRais Salsa MuhammadBelum ada peringkat
- Bioluminensi HewanDokumen12 halamanBioluminensi HewanPutri Aliyah100% (1)
- Ekstraksi DNADokumen19 halamanEkstraksi DNAcyrillakinanBelum ada peringkat
- Teori Fisiologi Mikroba "Microbial Stress Responses" by Bu Reno Fitri M.SiDokumen22 halamanTeori Fisiologi Mikroba "Microbial Stress Responses" by Bu Reno Fitri M.SiAzmi Azkiatul Fikri100% (1)
- Nukleus Dan NukleolusDokumen7 halamanNukleus Dan NukleolusAldhie Phoenix FarrellBelum ada peringkat
- Fungsi Dan Gambar AlatDokumen13 halamanFungsi Dan Gambar AlatFitra KhBelum ada peringkat
- Laporan FotosintesisDokumen7 halamanLaporan Fotosintesisdara intanBelum ada peringkat
- Preparat Squash Ujung Akar Bawang Merah (1,7,8,9)Dokumen11 halamanPreparat Squash Ujung Akar Bawang Merah (1,7,8,9)coklatklasikBelum ada peringkat
- Sisi Pemotongan Enzim Dan SequencingDokumen8 halamanSisi Pemotongan Enzim Dan SequencingNyda AnnidaaBelum ada peringkat
- Asam NukleatDokumen49 halamanAsam NukleathasnaBelum ada peringkat
- Makalah BioteknologiDokumen23 halamanMakalah Bioteknologi190384205058 Cilsa Nabila Hilal MandalikoBelum ada peringkat
- Makalah Ilmu Pengetahuan SosialDokumen13 halamanMakalah Ilmu Pengetahuan Sosialaisyahaisyahh94Belum ada peringkat
- Metode Maxam-GilbertDokumen28 halamanMetode Maxam-GilbertAnidraqif HastinaBelum ada peringkat
- Kerangka TeoriDokumen1 halamanKerangka TeoriAnidraqif HastinaBelum ada peringkat
- Bahan VirulensiDokumen19 halamanBahan VirulensiAnidraqif HastinaBelum ada peringkat
- Kerangka TeoriDokumen1 halamanKerangka TeoriAnidraqif HastinaBelum ada peringkat
- M. Amhar Jamil - Faktor Virulensi MikrobiologiDokumen23 halamanM. Amhar Jamil - Faktor Virulensi MikrobiologiAnidraqif HastinaBelum ada peringkat
- Transkripsi DNADokumen27 halamanTranskripsi DNAAnidraqif HastinaBelum ada peringkat
- EukariotDokumen25 halamanEukariotAnidraqif HastinaBelum ada peringkat
- Transkripsi DNADokumen27 halamanTranskripsi DNAAnidraqif HastinaBelum ada peringkat
- Buku Ajar Gen 10Dokumen15 halamanBuku Ajar Gen 10Anidraqif HastinaBelum ada peringkat
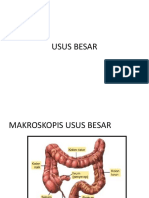
- M. Amhar Jamil - Usus Besar Dan Hormon GiDokumen7 halamanM. Amhar Jamil - Usus Besar Dan Hormon GiAnidraqif HastinaBelum ada peringkat
- Pelayan Informasi Obat Andi Ulfiana Utari n012181011Dokumen22 halamanPelayan Informasi Obat Andi Ulfiana Utari n012181011Anidraqif HastinaBelum ada peringkat
- M. Amhar Jamil - Faktor Virulensi MikrobiologiDokumen23 halamanM. Amhar Jamil - Faktor Virulensi MikrobiologiAnidraqif HastinaBelum ada peringkat
- Tugas Biomol 1 M. Amhar JamilDokumen17 halamanTugas Biomol 1 M. Amhar JamilAnidraqif HastinaBelum ada peringkat
- Tugas Biomol 1 M. Amhar JamilDokumen17 halamanTugas Biomol 1 M. Amhar JamilAnidraqif HastinaBelum ada peringkat
- Ekspresi GenDokumen28 halamanEkspresi GenYoudhystira Putra Asmara TanusasmitaBelum ada peringkat
- M. Amhar Jamil - Usus Besar Dan Hormon GiDokumen7 halamanM. Amhar Jamil - Usus Besar Dan Hormon GiAnidraqif HastinaBelum ada peringkat
- Laporan PKL PT. Phapros Tbk. SISTEM INSTDokumen115 halamanLaporan PKL PT. Phapros Tbk. SISTEM INSTAnidraqif HastinaBelum ada peringkat
- Imro Atul HasanaDokumen50 halamanImro Atul HasanaAnidraqif HastinaBelum ada peringkat
- SURVEI KEPUASAN PELANGGANDokumen62 halamanSURVEI KEPUASAN PELANGGANAnidraqif HastinaBelum ada peringkat
- LaporanDokumen7 halamanLaporanAnidraqif HastinaBelum ada peringkat
- Tugas Biomol 1 M. Amhar JamilDokumen22 halamanTugas Biomol 1 M. Amhar JamilAnidraqif HastinaBelum ada peringkat
- For WiwiDokumen10 halamanFor WiwiAnidraqif HastinaBelum ada peringkat
- Imro Atul HasanaDokumen50 halamanImro Atul HasanaAnidraqif HastinaBelum ada peringkat
- Bab IiDokumen50 halamanBab IiAnidraqif HastinaBelum ada peringkat
- Daftar TabelDokumen50 halamanDaftar TabelAnidraqif HastinaBelum ada peringkat
- Ujian Sidang ApotekDokumen12 halamanUjian Sidang Apotekabdillah amirBelum ada peringkat
- New Microsoft Office PowerPoint PresentationDokumen10 halamanNew Microsoft Office PowerPoint PresentationAnidraqif HastinaBelum ada peringkat
- PKPA ApotekDokumen55 halamanPKPA ApotekAnidraqif HastinaBelum ada peringkat
- Nasional CBTDokumen41 halamanNasional CBTnee_maneeeBelum ada peringkat