Insulin and Insulin Resistance
Diunggah oleh
Khansa RosviJudul Asli
Hak Cipta
Format Tersedia
Bagikan dokumen Ini
Apakah menurut Anda dokumen ini bermanfaat?
Apakah konten ini tidak pantas?
Laporkan Dokumen IniHak Cipta:
Format Tersedia
Insulin and Insulin Resistance
Diunggah oleh
Khansa RosviHak Cipta:
Format Tersedia
Machine Translated by Google
Lihat diskusi, statistik, dan profil penulis untuk publikasi ini di: https://www.researchgate.net/publication/7489006
Insulin dan Resistensi Insulin
Artikel dalam Ahli biokimia klinis. Ulasan / Asosiasi Ahli Biokimia Klinis Australia Juni 2005
Sumber: PubMed
KUTIPAN BACA
1.157 4.970
1 penulis:
Gisela Wilcox
Kepercayaan Yayasan Salford Royal NHS
68 PUBLIKASI 3.179 KUTIPAN
LIHAT PROFIL
Beberapa penulis publikasi ini juga mengerjakan proyek terkait berikut:
proyek CalciumView
Kesehatan mental dalam proyek PKU View
Semua konten setelah halaman ini diunggah oleh Gisela Wilcox pada tanggal 20 Mei 2014.
Pengguna telah meminta peningkatan file yang diunduh.
Machine Translated by Google
Mengulas artikel
Insulin dan Resistensi Insulin
Gisela Wilcox
Melbourne Pathology, Collingwood, VIC 3066, Monash University Department of Medicine & Clinical Nutrition & Metabolism
Unit, C/- Body Composition Laboratory, Monash Medical Centre, Clayton, VIC 3168, Australia
Untuk korespondensi: Dr Gisela Wilcox email: gisela .wilcox @med.monash.edu.au
Abstrak
Ketika obesitas dan diabetes mencapai proporsi epidemi di negara maju, peran resistensi insulin dan konsekuensinya semakin menonjol.
Memahami peran insulin dalam proses fisiologis yang luas dan pengaruhnya pada sintesis dan sekresinya, di samping tindakannya dari tingkat
molekuler hingga seluruh tubuh, memiliki implikasi signifikan untuk banyak penyakit kronis yang terlihat pada populasi kebarat-baratan saat ini.
Tinjauan ini memberikan ikhtisar tentang insulin, sejarahnya, struktur, sintesis, sekresi, aksi dan interaksinya diikuti dengan diskusi tentang
resistensi insulin dan manifestasi klinis terkaitnya. Area fokus khusus termasuk aksi insulin dan manifestasi resistensi insulin pada organ dan
jaringan tertentu, pengaruh fisiologis, lingkungan dan farmakologis terhadap aksi insulin dan resistensi insulin serta sindrom klinis yang terkait
dengan resistensi insulin. Langkah-langkah klinis dan fungsional resistensi insulin juga dibahas.
Terlepas dari pemahaman kita yang tidak lengkap tentang mekanisme biologis yang kompleks dari aksi insulin dan resistensi insulin, kita perlu
mempertimbangkan perubahan sosial yang dramatis di abad yang lalu sehubungan dengan aktivitas fisik, pola makan, pekerjaan, sosialisasi, dan
pola tidur. Globalisasi yang cepat, urbanisasi, dan industrialisasi telah melahirkan epidemi obesitas, diabetes, dan penyakit penyerta yang
menyertainya, karena ketidakaktifan fisik dan ketidakseimbangan pola makan membuka tabir sifat genetik predisposisi laten.
Perkenalan tidak akan lengkap, tetapi akan menetapkan adegan, menyatukan
Dengan obesitas dan diabetes yang mencapai proporsi epidemik di beragam bidang pemahaman saat ini tentang bidang yang terus
negara maju,1 peran resistensi insulin dan sekuelenya semakin muncul ini.
menonjol. Memahami peran insulin dalam berbagai proses fisiologis
dan pengaruhnya terhadap sintesis dan sekresinya, di samping Definisi dan Konsep Insulin
aksinya dari tingkat molekuler hingga seluruh tubuh, memiliki implikasi adalah hormon peptida yang disekresikan oleh sel ÿ pulau Langerhans
yang signifikan untuk banyak penyakit kronis yang terlihat pada pankreas dan mempertahankan kadar glukosa darah normal dengan
populasi kebarat-baratan saat ini. Akibatnya, lebih dari satu abad memfasilitasi pengambilan glukosa seluler, mengatur metabolisme
setelah para ilmuwan mulai menjelaskan peran pankreas pada karbohidrat, lipid dan protein dan mendorong pembelahan dan
diabetes, studi tentang insulin dan resistensi insulin tetap menjadi pertumbuhan sel melalui efek mitogeniknya.
yang terdepan dalam penelitian medis, relevan di semua tingkatan
dari bangku ke samping tempat tidur dan kebijakan kesehatan
masyarakat. Resistensi insulin didefinisikan di mana insulin normal atau meningkat
tingkat menghasilkan respon biologis dilemahkan;2 klasik ini mengacu
Lingkup Tinjauan pada sensitivitas gangguan terhadap insulin dimediasi glukosa
Tinjauan ini akan mencakup ikhtisar insulin, riwayatnya, struktur, pembuangan.3
sintesis, sekresi, aksi dan interaksinya diikuti dengan diskusi tentang
resistensi insulin dan manifestasi klinis terkaitnya. Bagian terakhir ini Hiperinsulinemia kompensasi terjadi ketika sekresi sel ÿ pankreas
akan meningkat untuk mempertahankan kadar glukosa darah normal dalam
mendiskusikan mekanisme dan konteks, fisiologis dan patologis, pengaturan resistensi insulin perifer di otot dan jaringan adiposa.
resistensi insulin, dan penilaiannya di laboratorium klinis. Mengingat
luasnya topik, ulasan ini
Klinik Biochem Rev Vol 26 Mei 2005 I 19
Machine Translated by Google
WilcoxG
Sindrom resistensi insulin mengacu pada sekelompok kelainan dan Struktur dan Sifat Kimia Insulin Insulin ditemukan
hasil fisik terkait yang lebih sering terjadi pada individu yang resistan sebagai polipeptida pada tahun 1928 dengan rangkaian asam
terhadap insulin. Mengingat perbedaan jaringan dalam aminonya diidentifikasi pada tahun 1952. Insulin sebenarnya
ketergantungan dan sensitivitas insulin, manifestasi dari sindrom merupakan dipeptida, yang masing-masing mengandung rantai A
resistensi insulin cenderung mencerminkan efek gabungan dari dan B, dihubungkan oleh jembatan disulfida, dan mengandung 51
kelebihan insulin dan resistensi variabel terhadap aksinya.3 asam amino, dengan berat molekul 5802. Titik iso-listriknya adalah
pH 5.5.5 Rantai A terdiri dari 21 asam amino dan rantai B 30 asam
amino. Rantai A memiliki heliks terminal-N yang terhubung ke
Sindrom metabolik mewakili entitas diagnostik klinis yang heliks terminal C anti-paralel; rantai B memiliki segmen heliks pusat.
mengidentifikasi orang-orang yang berisiko tinggi sehubungan Kedua rantai dihubungkan oleh 2 ikatan disulfida, yang
dengan morbiditas (kardiovaskular) yang terkait dengan resistensi menghubungkan heliks terminal-N dan C dari rantai A ke heliks pusat.
insulin.3 atau Bchain. Dalam proinsulin, peptida penghubung menghubungkan
ujung-N rantai A ke ujung-C rantai B.6
Penemuan Insulin Pada
tahun 1889 ilmuwan Jerman Minkowski dan von Mering mencatat, Sintesis dan Pelepasan Insulin
dari percobaan mereka dengan hewan, bahwa pankreatektomi total Insulin dikodekan pada lengan pendek kromosom 117 dan disintesis
menyebabkan perkembangan diabetes parah.4 Mereka berhipotesis dalam sel ÿ pulau pankreas Langherhans sebagai prekursornya,
bahwa zat yang dikeluarkan oleh pankreas bertanggung jawab proinsulin. Proinsulin disintesis di ribosom retikulum endoplasma
untuk pengendalian metabolisme. Yang lain kemudian kasar (RER) dari mRNA sebagai pra-proinsulin. Pre-proinsulin
menyempurnakan hipotesis ini, mencatat bahwa diabetes dikaitkan dibentuk oleh sintesis berurutan dari peptida sinyal, rantai B, peptida
dengan penghancuran pulau Langerhans. Sementara Minkowski, penghubung (C) dan kemudian rantai A yang terdiri dari rantai
serta Zuelzer di Jerman dan Scott di Amerika Serikat berusaha, tunggal dari 100 asam amino. Penghapusan peptida sinyal
dengan hasil yang tidak konsisten, untuk mengisolasi dan mengelola membentuk proinsulin, yang memperoleh struktur 3 dimensi khasnya
substansi pulau pankreas yang hilang, penyelidik Belgia de Meyer di retikulum endoplasma. Vesikel sekretorik mentransfer proinsulin
pada tahun 1909 mengusulkan nama "insulin", seperti yang dari RER ke aparatus Golgi, yang lingkungan kaya seng dan
dilakukan peneliti Inggris Schaefer pada tahun 1916. . kalsiumnya mendukung pembentukan hexamers proinsulin yang
mengandung seng.6 Saat vesikel penyimpanan yang belum matang
Akhirnya pada tahun 1921, satu dekade kemudian, insulin akhirnya terbentuk dari Golgi, enzim yang bekerja di luar Golgi mengubah
diisolasi, dimurnikan, dan tersedia dalam bentuk yang dapat proinsulin menjadi insulin dan C peptide.8 Insulin membentuk
digunakan untuk terapi. Pada Mei 1921, ahli bedah Toronto Banting, heksamer yang mengandung seng yang tidak larut, mengendap
dibantu oleh mahasiswa kedokteran Best, dan di bawah pengawasan sebagai kristal yang stabil secara kimiawi pada pH 5,5. Ketika
McLeod, Profesor Metabolisme Karbohidrat, memulai percobaan butiran matang disekresikan ke dalam sirkulasi oleh eksositosis,
pada anjing. Mereka memberikan ekstrak garam pankreas dingin insulin, dan rasio equimolar C-peptida dilepaskan. Proinsulin dan
secara intravena ke anjing yang mengalami diabetes dengan seng biasanya terdiri tidak lebih dari 6% dari sekresi sel islet.6
pankreatektomi dan mengamati penurunan glukosa darah.
Pada bulan Desember 1921, karya ini dipresentasikan kepada
American Physiological Association, dan ahli biokimia Collip, yang
telah bergabung dengan tim tersebut, selanjutnya menunjukkan Sekresi insulin dari sel-sel islet ke dalam vena portal secara khas
bahwa ekstrak ini juga mengembalikan mobilisasi glikogen hati dan berdenyut, mencerminkan penjumlahan semburan sekresi koordinat
kapasitas untuk membersihkan keton. Satu bulan kemudian, pada dari jutaan sel islet. Sebuah pola ultradian osilasi pelepasan insulin,
Januari 1922, percobaan manusia pertama dimulai pada seorang selain variasi setelah makan, telah dilaporkan.9 Menanggapi
anak laki-laki penderita diabetes berusia 14 tahun yang gejala klinis stimulus seperti glukosa, sekresi insulin bersifat bifasik, dengan
dan kelainan biokimianya pada dasarnya dibalik dengan pemberian fase awal sekresi insulin yang cepat, diikuti dengan fase yang
isolat pankreas. Pada bulan Mei 1922, komponen aktif tersebut kurang intens tetapi pelepasan hormon yang lebih berkelanjutan.10
diberi nama insulin, dan hasil percobaan ini dipresentasikan kepada
Asosiasi Dokter Amerika. Eli Lilly kemudian memulai produksi insulin
babi, meningkatkan pemurnian melalui presipitasi iso-listrik,
menghasilkan jumlah komersial pada awal 1923. Hadiah Nobel Faktor-Faktor yang Mempengaruhi Biosintesis dan
diberikan pada 1923 kepada Banting dan McLeod.4 Pelepasan Insulin Sekresi insulin dapat dipengaruhi oleh perubahan
sintesis pada tingkat transkripsi gen, translasi, dan modifikasi pasca-
translasi di Golgi serta oleh faktor-faktor yang mempengaruhi
pelepasan insulin dari butiran sekretori.
20 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
Modifikasi jangka panjang dapat terjadi melalui pengaruh pada massa dalam sekresi insulin pankreas. Hal ini dianggap memediasi apa yang
dan diferensiasi sel ÿ.11 Mengingat peran penting insulin dalam disebut "fase cephalic" sekresi insulin, yang terjadi saat makanan
pemanfaatan dan metabolisme glukosa, tidak mengherankan bahwa dilihat, dicium, atau ditelan secara akut. Reseptor muskarinik kolinergik
glukosa memiliki banyak pengaruh pada biosintesis dan sekresi insulin. sel pulau mengaktifkan fosfolipase C, dengan kejadian intraseluler
Namun, faktor lain seperti asam amino, asam lemak, asetilkolin, selanjutnya mengaktifkan protein kinase C, fosfolipase A2 dan
polipeptida pengaktif adenilat siklase hipofisis (PACAP), polipeptida memobilisasi kalsium intraseluler.
insulinotropik yang bergantung pada glukosa (GIP), glukagon-like Sekresi insulin melalui mekanisme ini tidak terjadi pada keadaan puasa
peptide-1 (GLP-1), dan beberapa agonis lainnya, bersama-sama dalam atau jika kadar glukosa darah rendah, tetapi dapat meningkatkan
kombinasi, juga mempengaruhi proses ini.10 respons anabolik terhadap pemberian makan.8
2. Jalur Adrenergik
Mekanisme Sekresi Insulin Katekolamin, melalui ÿ2-adrenoseptor, biasanya menghambat
Peningkatan kadar glukosa menginduksi "fase pertama" sekresi insulin pelepasan insulin selama stres dan olahraga. Pankreas juga memiliki
yang dimediasi glukosa dengan pelepasan insulin dari butiran sekretori ÿ-adrenoseptor, yang meningkatkan sekresi insulin melalui cAMP,
dalam sel ÿ. Entri glukosa ke dalam sel ÿ dirasakan oleh glukokinase, tetapi efek ini tampaknya kecil. Efek yang dimediasi ÿ2 tampaknya
yang memfosforilasi glukosa menjadi glukosa-6-fosfat (G6P), bertindak hilir dari efek secretogogues nutrisi.8
menghasilkan ATP.12 Penutupan saluran yang bergantung pada K+
-ATP menghasilkan depolarisasi dan aktivasi dalam membran Hormon Peptida
saluran kalsium yang bergantung pada voltase yang menyebabkan Sejumlah hormon peptida mempengaruhi sekresi insulin. Hormon usus
peningkatan konsentrasi kalsium intraseluler; hal ini memicu sekresi GLP-114 dan somatostatin masing-masing meningkatkan dan
insulin pulsatil. Peningkatan respons ini terjadi baik oleh jalur K+ -ATP menghambat kerja insulin. Sekresi mereka dirangsang oleh nutrisi di
yang bergantung pada saluran Ca2+ dan jalur yang tidak bergantung usus. Mekanisme aksi mereka tampaknya, setidaknya sebagian,
pada saluran K+ -ATP Ca2+ yang tidak tergantung saluran dari aksi dimediasi oleh pembentukan cAMP dan aktivasi protein kinase yang
glukosa. Mediator lain dari pelepasan insulin termasuk aktivasi responsif terhadap cAMP.8 Leptin dan adiponektin tampaknya bekerja
dengan cara yang serupa sedangkan protein penstimulasi asilasi
fosfolipase dan protein kinase C (misalnya dengan asetilkolin) dan tampaknya bekerja melalui fosforilasi glukosa, kalsium influ ux dan
dengan stimulasi aktivitas adenilil siklase dan aktivasi protein kinase A protein kinase C.15
sel ÿ, yang mempotensiasi sekresi insulin.
Mekanisme terakhir ini dapat diaktifkan oleh hormon, seperti peptida Asam amino
usus vasoaktif (VIP), PACAP, GLP-1, dan GIP. Efek insulinotropik dari arginin asam amino kationik sudah mapan. L-
Faktor-faktor ini tampaknya memainkan peran penting dalam fase ornithine memiliki efek yang serupa. Tindakan arginin dikaitkan dengan
kedua sekresi insulin yang dimediasi glukosa, setelah pengisian kembali peningkatan permeabilitas kalium tanpa efek apa pun pada biosintesis
granula sekretori yang ditranslokasi dari kumpulan cadangan.10 proinsulin. Penyerapan arginin ke dalam sel ÿ dikaitkan dengan
depolarisasi membran sel dan gerbang saluran kalsium yang bergantung
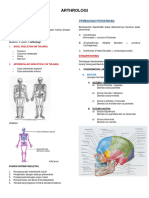
Regulasi dan Mekanisme Sekresi Insulin di pada tegangan.8 Leusin, asam amino rantai cabang esensial, adalah
Tingkat seluler peningkat kuat sekresi insulin, merangsang glutamat dehidrogenase
Sintesis dan sekresi insulin diatur oleh secretagog nutrisi dan non- secara alosterik dan membentuk ÿ-ketoglutarat melalui siklus Krebs
nutrien, dalam konteks rangsangan lingkungan dan interaksi hormon untuk menghasilkan ATP.12 Jalur utama dan pengaruh pada sekresi
lain.8 Sekretagog nutrisi seperti glukosa tampaknya memicu sekresi insulin disajikan secara skematis pada Gambar 1.
insulin dari sel ÿ dengan meningkatkan ATP intraseluler dan penutupan
atau saluran K+ -ATP seperti diuraikan di atas. Generasi AMP siklik
dan perantara energi seluler lainnya juga ditambah, semakin
meningkatkan pelepasan insulin. Glukosa tidak memerlukan aksi insulin Faktor-Faktor yang Mempengaruhi
untuk memasuki sel ÿ (tidak juga fruktosa, mannosa atau galaktosa). Massa Sel ÿ Regulasi sekresi insulin jangka panjang dapat dimediasi
Sekretagog non-nutrisi dapat bekerja melalui rangsangan saraf seperti melalui efek pada massa sel ÿ. Hormon pertumbuhan, prolaktin,
jalur kolinergik dan adrenergik, atau melalui hormon peptida dan asam laktogen plasenta, dan GLP-1 tidak hanya meningkatkan pelepasan
amino kationik. insulin terstimulasi glukosa dan ekspresi gen insulin, tetapi juga
meningkatkan proliferasi sel ÿ.11,14 Untuk tinjauan komprehensif
tentang topik ini lihat Nielsen dan Serup.16
Stimulus saraf
1. Transmisi Kolinergik Telah Fisiologi Sekresi Insulin
diketahui dengan baik bahwa hasil stimulasi saraf vagus Glukosa adalah stimulus utama untuk sekresi insulin
Klinik Biochem Rev Vol 26 Mei 2005 I 21
Machine Translated by Google
WilcoxG
Gambar 1. Skema presentasi jalur sekresi insulin. Diadaptasi dari referensi: 10 & 14.
makronutrien lain, hormon, faktor humoral, dan masukan saraf konsentrasi insulin puasa vena (atau arteri) yang bersirkulasi
dapat mengubah respons ini. Insulin, bersama dengan glukagon adalah sekitar 3-15 mIU/L atau 18-90 pmol/L.17 Sekresi insulin
hormon kontra-regulasi utamanya, mengatur konsentrasi terkait makanan merupakan fraksi yang tersisa dari total output
glukosa darah.17 Sel ÿ pankreas mengeluarkan 0,25-1,5 unit harian.
insulin per jam selama keadaan puasa (atau basal), cukup
untuk mengaktifkan glukosa yang bergantung pada insulin Sekresi Insulin dalam Menanggapi
masuk ke dalam sel. Tingkat ini mencegah hidrolisis trigliserida Rangsangan
yang tidak terkendali dan membatasi glukoneogenesis, Tanggapan terhadap Glukosa Pada orang sehat, sekresi
sehingga mempertahankan kadar glukosa darah puasa normal. pankreas yang distimulasi glukosa bersifat bifasik. Pemberian
Sekresi insulin basal menyumbang lebih dari 50% dari total sekresi insulin
glukosa24secara
jam. intravena dikaitkan dengan “fase pertama”
Mengikuti sekresi insulin ke dalam sistem vena portal, 60% pelepasan insulin yang cepat dalam 1 menit, memuncak pada
selanjutnya dibuang oleh hati; sehingga konsentrasi insulin 3-5 menit, dan berlangsung sekitar 10 menit; onset lambat
vena porta yang mencapai hati mendekati tiga kali lipat dari "fase kedua" sekresi insulin dimulai segera setelah bolus
sirkulasi perifer. Pada individu kurus yang sehat glukosa tetapi tidak terlihat sampai 10 menit kemudian, berlangsung duras
Catatan kaki: Singkatan Gambar
Akt/PKB = protein kinase B GLP-1 = peptida 1 mirip glukagon
ATP = adenosin trifosfat IRS = substrat reseptor insulin
ADP = adenosin difosfat cAMP = MAP kinase = protein kinase teraktivasi mitogen
adenosin monofosfat siklik PACAP = polipeptida pengaktif adenilat siklase hipofisis
HARI = diasilgliserol PI3K = fosfatidilinositol 3-kinase
GIP = peptida penghambat lambung / PIPD1 & 2 = protein kinase 1 & 2 yang bergantung pada fosfatidilinositol
polipeptida insulinotropik yang bergantung pada glukosa PKC = protein kinase C
Glukosa- 6- P = glukosa 6 fosfat RAS = protein sarkoma tikus
GLUT 2 = protein pengangkut glukosa 2 SHC = protein adaptor dengan homologi src
GLUT 4 = protein pengangkut glukosa 4 VIP = peptida usus vasoaktif
22 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
sebanding dengan konsentrasi glukosa sesaat sebelum pemberian hormon inkretin yang paling penting. GLP-1 juga menghambat pelepasan
glukosa.17 Fase pertama sekresi insulin merupakan pelepasan insulin glukagon, menunda pengosongan lambung dan mengurangi nafsu makan.24
yang sudah disintesis dan disimpan dalam butiran sekretori; fase
kedua mewakili sekresi insulin yang disimpan dan yang baru disintesis. Pengaruh Rangsangan Syaraf dan Hormonal
Sekresi insulin secara keseluruhan berhubungan dengan dosis total Lihat Tabel 1.
glukosa dan kecepatan pemberiannya; respon pankreas maksimal
terjadi dengan 20 g glukosa diberikan secara intravena selama 3 menit
Reseptor Insulin dan Pengikatan Insulin
pada manusia.18
Insulin memediasi aksinya melalui pengikatan pada reseptor insulin.
Reseptor insulin pertama kali dikarakterisasi pada tahun 1971. Ini
terdiri dari heterotetramer yang terdiri dari 2 subunit glikoprotein ÿ dan
Berbeda dengan pola sekresi insulin yang dapat direproduksi sebagai
2 ÿ yang dihubungkan oleh ikatan disulfida dan terletak di membran
respons terhadap glukosa intravena, sekresi insulin setelah glukosa
sel.25 Gen yang mengkode reseptor insulin terletak di lengan pendek
oral jauh lebih bervariasi. Dengan beban glukosa oral, pengosongan
reseptor insulin . kromosom 19.17 Insulin berikatan dengan subunit ÿ
lambung dan motilitas gastrointestinal mempengaruhi penyerapan
ekstraseluler, menghasilkan perubahan konformasi yang
glukosa, hormon gastrointestinal dan masukan saraf yang terkait
memungkinkan ATP berikatan dengan komponen intraseluler subunit
dengan konsumsi glukosa memodifikasi respon insulin, dan sekresi
ÿ.23 Pengikatan ATP pada gilirannya memicu fosforilasi subunit ÿ
insulin berlanjut beberapa saat setelah konsumsi glukosa.17
yang memberikan aktivitas tirosin kinase. Hal ini memungkinkan
fosforilasi tirosin protein substrat intraseluler yang dikenal sebagai
substrat responsif insulin (IRS). IRS kemudian dapat mengikat molekul
Respon terhadap Arginin
pensinyalan lain yang memediasi tindakan seluler lebih lanjut dari
Respon terhadap arginin ditandai dengan baik. Sekresi insulin yang insulin.25
diinduksi arginin sebanding dengan kadar glukosa basal dan terjadi
2-10 menit setelah injeksi IV.17 Konsumsi asam amino oral
Ada empat protein IRS bernama khusus yang diketahui. IRS 1 dan 2
merangsang sekresi insulin, serta hormon pertumbuhan dan
memiliki distribusi jaringan yang tumpang tindih. IRS 1 difosforilasi
glukagon.19
oleh reseptor insulin dan insulin like growth factor 1 (IGF-1 lihat di
bawah) reseptor, memediasi efek mitogenik insulin dan penginderaan
Efek Lipid Dalam
glukosa pasangan terhadap sekresi insulin dengan IRS 1 diusulkan
sirkulasi, asam lemak non-esterifikasi (NEFA) dapat berasal dari lipid
menjadi IRS utama dalam otot rangka. IRS 2, diusulkan untuk menjadi
makanan dan dilepaskan oleh hati, atau disintesis dalam adiposit, hati,
IRS utama di hati, memediasi aksi insulin perifer dan pertumbuhan sel
kulit dan usus kecil dari kelebihan karbohidrat. NEFA tidak hanya
ÿ pankreas.25 IRS 3 dan 4 memiliki karakteristik yang kurang baik.
meningkatkan keluaran glukosa hati dan mengurangi sensitivitas
IRS 3 hanya ditemukan di jaringan adiposa, sel ÿ dan hati dan IRS 4
insulin perifer, tetapi juga dapat memodifikasi sekresi insulin yang
di timus, otak dan ginjal.26,27 Protein IRS terfosforilasi mengikat
distimulasi glukosa (GSIS). Peningkatan akut NEFA plasma dikaitkan
protein domain src-homology-2 spesifik (SH2), yang termasuk enzim
dengan peningkatan GSIS, tergantung pada konsentrasi glukosa yang
penting seperti phosphatidylinositol 3 -kinase (PI 3-kinase) dan
ada.
Peningkatan kronis NEFA dikaitkan dengan penurunan phosphotyrosine phosphatase SHPTP2 (atau Syp), dan protein lain
yang tidak memiliki aktivitas enzimatik tetapi menghubungkan IRS-1
di GSIS, dan berkurangnya sintesis insulin.20 Penelanan lipid juga
dan sistem pensinyalan intraseluler lainnya, misalnya protein adaptor
dapat mengubah respons insulin terhadap glukosa melalui efek pada
Grb2 yang terhubung dengan RAS (protein sarkoma tikus ) jalan.
hormon gastrointestinal dan pengosongan lambung.21
Respons terhadap Makanan
Campuran Ini adalah rangsangan yang jauh lebih fisiologis untuk
sekresi insulin. Pola sekresi insulin akan bergantung pada proporsi PI 3-kinase mendorong translokasi protein pengangkut glukosa,
relatif berbagai nutrisi, bentuk fisik komponen makanan yang dicerna sintesis glikogen, lipid dan protein, anti-lipolisis, dan kontrol
dan faktor-faktor yang mempengaruhi respons terhadap glukosa oral glukoneogenesis hepatik.27 PI 3-kinase bekerja melalui serin dan
saja.22,23 treonin kinase seperti Akt/protein kinase B (PKB) , protein kinase C
(PKC) dan protein kinase1 & 2 yang bergantung pada PI (PIPD 1 &
Hormon Inkretin 2). Jalur RAS mengaktifkan faktor transkripsi dan menstimulasi aksi
Nutrisi dalam saluran GI merangsang sekresi hormon peningkatan pertumbuhan insulin.25 Dengan demikian secara luas, PI
dikenal sebagai incretins yang memperkuat pelepasan insulin yang 3-kinase memediasi efek metabolik insulin, misalnya penyerapan
diinduksi glukosa. Ini menyebabkan respons insulin yang lebih besar glukosa seluler, sementara RAS secara signifikan memediasi efek
terhadap glukosa oral, bukan intravena. GIP dan GLP-1 adalah keduanya mitogenik insulin17,25
Klinik Biochem Rev Vol 26 Mei 2005 I 23
Machine Translated by Google
WilcoxG
Tabel 1. Mediator sekresi insulin.
rangsangan Gizi Hormon saraf
Stimulasi Glukosa hormon pertumbuhan ÿ-adrenergik
asam amino Glukagon Vagal
(keton) GLP-1 (parasimpatis)
BOHONG
Rahasia
Kolesistokinin
Gastrin
VIP
Peptida pelepas gastrin
Penghambatan Adrenocorticosteroid ÿ-adrenergik
Somatostatin
adrenalin
Norepinefrin
Galanin
Neuropeptida Y
Peptida terkait gen kalsitonin
(CGRP)
Prostaglandin E
Referensi: Diadaptasi dari referensi 17.
bersama-sama dengan tindakan lain yang kurang dijelaskan dengan tertindas. Namun, ketika kadar glukosa dan insulin turun ke nilai
baik. Jalur ini disajikan secara skematis pada Gambar 2. puasa, glukosa tidak lagi masuk ke dalam sel, mendorong lipolisis.
Dalam sel otot, transpor glukosa intraseluler memfasilitasi sintesis
Protein Pengangkut Glukosa glikogen dalam keadaan makan.27 PI 3-kinase tampaknya penting
Glukosa memasuki sel dengan cara yang tidak tergantung ATP untuk translokasi GLUT 4 ke membran sel dalam sel otot dan
melalui protein pengangkut glukosa (GLUT), di mana setidaknya 5 adiposit; ini memfasilitasi aksi hilir dari enzim kunci intraseluler ini.25
subtipe telah diidentifikasi28,29 ( Tabel 2). Berbeda dalam karakteristik
seperti Km untuk transportasi glukosa maksimal dan ketergantungan
insulin, mereka memungkinkan jenis sel yang berbeda untuk Tindakan Insulin di Tingkat Seluler
memanfaatkan glukosa sesuai dengan fungsi spesifiknya. Misalnya, Tindakan insulin pada tingkat sel meliputi metabolisme karbohidrat,
sebagian besar sel otak, yang memiliki GLUT 1 sebagai protein lipid dan asam amino serta transkripsi dan translasi mRNA.
pengangkut utama, mampu memindahkan glukosa secara intraseluler
pada konsentrasi glukosa darah yang sangat rendah tanpa
membutuhkan insulin. Jadi neuron-neuron ini, yang pada prinsipnya Metabolisme Karbohidrat
bergantung pada glukosa untuk energi intraseluler, mampu Insulin bekerja pada beberapa langkah dalam metabolisme
mengekstraknya dari sirkulasi dan berfungsi meskipun kadar glukosa karbohidrat. Efeknya pada difusi terfasilitasi glukosa menjadi lemak
dan insulin rendah terlihat selama keadaan puasa. Di sisi lain, sel dan sel otot melalui modulasi translokasi GLUT 4 telah terjadi
adiposa dan sel otot memiliki GLUT 4 sebagai protein pengangkut dibahas. Sintesis glikogen meningkat, dan pemecahan glikogen
glukosa utama, yang membutuhkan insulin untuk kerjanya dan menurun, masing-masing dengan defosforilasi glikogen sintase dan
memiliki Km yang jauh lebih tinggi untuk glukosa. Hal ini glikogen fosforilase kinase.
memungkinkan sel-sel jaringan adiposa, yang fungsinya untuk Glikolisis dirangsang dan glukoneogenesis dihambat oleh defosforilasi
menyimpan energi berlebih, untuk merespons karakteristik kadar piruvat kinase (PK) dan 2,6 bifosfat kinase. Menandakan kelimpahan
glukosa yang lebih tinggi dari keadaan makan, dan memungkinkan energi intraseluler, insulin meningkatkan konversi piruvat menjadi
ireversibel
glukosa memasuki sel tempat sintesis asam lemak dan gliserol distimulasi dan lipolisis.
24 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
Gambar 2. Presentasi skema jalur pensinyalan insulin. Diadaptasi dari referensi: 25, 28 & 35. Lihat Catatan Kaki di halaman 22
untuk singkatan Gambar.
Acetyl Co-A dengan aktivasi kompleks enzim piruvat Sintesis Protein
dehidrogenase intra-mitokondria. Asetil-KoA kemudian dapat Insulin meningkatkan sintesis protein dalam berbagai jaringan.
langsung dioksidasi melalui siklus Krebs, atau digunakan untuk Ada efek pada transkripsi mRNA spesifik, serta translasi mRNA
sintesis asam lemak.30 menjadi protein di ribosom.
Contoh peningkatan transkripsi mRNA termasuk mRNA untuk
Metabolisme Lipid glukokinase, PK, sintase asam lemak dan albumin di hati,
Insulin merangsang sintesis asam lemak di jaringan adiposa, piruvat karboksilase di jaringan adiposa, kasein di kelenjar susu
hati dan kelenjar susu menyusui bersama dengan pembentukan dan amilase di pankreas. Aksi insulin menurunkan mRNA untuk
dan penyimpanan trigliserida di jaringan adiposa dan hati. enzim hati seperti karbamoil fosfat sintetase, enzim kunci dalam
Sintesis asam lemak ditingkatkan dengan aktivasi dan siklus urea. Efek pada translasi tersebar luas dan dipengaruhi
peningkatan fosforilasi asetil-KoA karboksilase, sedangkan oleh insulin itu sendiri dan berbagai faktor pertumbuhan,
oksidasi lemak ditekan dengan penghambatan karnitin misalnya IGF-1.19,28
asiltransferase. Sintesis trigliserida distimulasi oleh esterifikasi
gliserol fosfat, sedangkan pemecahan trigliserida ditekan oleh
defosforilasi lipase sensitif hormon. Sintesis kolesterol meningkat Ligan Lain untuk Reseptor Insulin Insulin-
dengan aktivasi dan defosforilasi reduktase HMG Co-A like growth factor (IGF) disebut demikian karena memiliki
sementara pemecahan kolesterol ester tampaknya dihambat homologi struktural yang signifikan dengan proinsulin tetapi
oleh defosforilasi kolesterol esterase. Metabolisme fosfolipid terutama efek mitogenik, secara signifikan diatur oleh hormon
juga dipengaruhi oleh insulin.28 pertumbuhan.31 IGF-1 dan 2 dikodekan pada paru-paru
lengan kromosom 12 dan lengan pendek kromosom 11
masing-masing.32 Mereka memiliki reseptor spesifik dan berikatan dengan
Klinik Biochem Rev Vol 26 Mei 2005 I 25
Machine Translated by Google
WilcoxG
Tabel 2. Protein transporter glukosa.26,27
Distribusi Jaringan ISOFORM Afinitas untuk km Karakteristik Gen
Glukosa lokasi
Kekenyangan 1 pembuluh mikro otak, Di mana-mana
Hemat sel darah Tinggi 1 mmol/L Chr 1
Plasenta pengangkut dasar
Ginjal
Semua jaringan
Kekenyangan 2 Hati Transporter km tinggi
sel ÿ Rendah 15-20 mmol/L Chr 3
ginjal Insulin independen
Usus halus
Kekenyangan 3 neuron otak Transporter Km Rendah
Plasenta Tinggi <1 mmol/L Chr 12
Otot janin Ditemukan di jaringan
Semua jaringan yang bergantung pada glukosa
Kekenyangan 4 Sel otot Diasingkan
Sel lemak Sedang 2,5-5 mmol/L secara intraseluler dan Chr 17
Jantung mentranslokasi ke
permukaan sel sebagai
respons terhadap insulin
Kekenyangan 5 Usus halus Afinitas tinggi untuk
Testis Sedang 6mmol/L fruktosa Chr 1
afinitas yang berbeda dengan berbagai protein pengikat IGF. Insulin dapat asam amino otot. Dengan adanya suplai asam amino yang cukup, insulin
berikatan dengan reseptor untuk IGF-1 dan 2 tetapi dengan afinitas yang bersifat anabolik di otot.29
jauh lebih rendah (10-2 dan 5x10-3) . IGF-1 berikatan lemah dengan
reseptor insulin, dengan hanya 1,25x10-3 afinitas untuk reseptor IGF-1; itu Mekanisme Resistensi Insulin
mengikat reseptor IGF-2 dengan 1/4 afinitas untuk reseptor IGF-2. IGF-2 Secara fisiologis, pada tingkat seluruh tubuh, kerja insulin dipengaruhi oleh
tidak berikatan dengan reseptor insulin; itu tidak mengikat reseptor IGF-1 interaksi hormon lain. Insulin, meskipun merupakan hormon dominan yang
tetapi dengan 1/3 afinitas untuk reseptor IGF-2. Oleh karena itu tumpang menggerakkan proses metabolisme dalam keadaan kenyang, bekerja
tindih dalam fungsi fisiologis lebih terbatas in vivo. bersama dengan hormon pertumbuhan dan IGF 1; hormon pertumbuhan
disekresikan sebagai respons terhadap insulin, di antara rangsangan
lainnya, mencegah hipoglikemia yang diinduksi insulin.
Peran Fisiologis Insulin Insulin Hormon kontra-regulasi lainnya termasuk glukagon, glukokortikoid, dan
adalah hormon penting yang mengatur pasokan energi sel dan katekolamin. Hormon-hormon ini mendorong proses metabolisme dalam
keseimbangan makronutrien, mengarahkan proses anabolik dari keadaan keadaan puasa. Glukagon mempromosikan glikogenolisis, glukoneogenesis
makan.27 Insulin sangat penting untuk transpor glukosa intraseluler ke dan ketogenesis. Rasio insulin terhadap glukagon menentukan derajat
jaringan yang bergantung pada insulin seperti otot dan jaringan adiposa. fosforilasi atau defosforilasi enzim yang relevan.29 Katekolamin mendorong
Menandakan kelimpahan energi eksogen, pemecahan lemak jaringan lipolisis dan glikogenolisis; glukokortikoid meningkatkan katabolisme otot,
adiposa ditekan dan sintesisnya dipromosikan. Dalam sel otot, masuknya glukoneogenesis dan lipolisis.
glukosa memungkinkan glikogen untuk disintesis dan disimpan, dan untuk
karbohidrat, daripada asam lemak (atau asam amino) untuk digunakan Kelebihan sekresi hormon ini dapat menyebabkan resistensi insulin pada
sebagai sumber energi yang tersedia segera untuk kontraksi otot. Oleh pengaturan tertentu, tetapi tidak memperhitungkan sebagian besar keadaan
karena itu, insulin meningkatkan sintesis glikogen dan lipid dalam sel otot, resistensi insulin.
sambil menekan lipolisis dan glukoneogenesis
26 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
Resistensi insulin dalam banyak kasus diyakini bermanifestasi di penyerapan glukosa, mempromosikan lipogenesis sambil menekan
tingkat sel melalui defek pasca-reseptor dalam pensinyalan insulin. lipolisis, dan karenanya aliran asam lemak bebas ke dalam aliran darah.
Meskipun temuan yang menjanjikan pada hewan percobaan Karena adiposit tidak bergantung pada glukosa dalam keadaan basal,
sehubungan dengan berbagai cacat pensinyalan insulin, relevansinya energi intraseluler dapat disuplai oleh oksidasi asam lemak pada
dengan resistensi insulin manusia saat ini tidak jelas. Mekanisme keadaan defisiensi insulin, sambil melepaskan asam lemak bebas ke
yang mungkin termasuk down-regulation, defisiensi atau polimorfisme dalam sirkulasi untuk penggunaan langsung oleh organ lain misalnya
genetik fosforilasi tirosin dari reseptor insulin, protein IRS atau PIP-3 jantung, atau hati. di mana mereka diubah menjadi badan keton.
kinase, atau mungkin melibatkan kelainan fungsi GLUT 4.33 Badan keton menyediakan substrat energi alternatif untuk otak selama
kelaparan berkepanjangan.30
Situs Aksi Insulin dan Manifestasi Insulin Pada resistensi insulin, efeknya pada jaringan adiposa serupa, tetapi
Perlawanan di hati peningkatan fluks asam lemak bebas cenderung mendorong
Efek insulin, defisiensi insulin, dan resistensi insulin bervariasi sesuai produksi lipoprotein densitas sangat rendah (VLDL) hati37 sementara
dengan fungsi fisiologis jaringan dan organ yang bersangkutan, dan ketogenesis biasanya tetap ditekan oleh hiperinsulinemia kompensasi.
ketergantungannya pada insulin untuk proses metabolisme. Jaringan- Selain itu, karena aktivitas lipoprotein lipase bergantung pada insulin
jaringan yang didefinisikan sebagai ketergantungan insulin, dan terganggu oleh resistensi insulin, penyerapan trigliserida perifer
berdasarkan transpor glukosa intraseluler, pada dasarnya adalah dari VLDL juga berkurang. Mekanisme ini berkontribusi pada
jaringan adiposa dan otot. Namun, aksi insulin bersifat pleotropik dan
tersebar luas, demikian pula manifestasi resistensi insulin dan hipertrigliseridemia yang diamati dari resistensi insulin.38 Selain asam
hiperinsulinemia kompensasi terkait.3 lemak bebas, jaringan adiposa mengeluarkan sejumlah sitokin yang
memiliki efek sistemik pada resistensi insulin.
otot Ini termasuk IL-6, TNFÿ, plasminogen activator inhibitor 1 (PAI-1),
Penyerapan glukosa ke dalam otot pada dasarnya bergantung pada angiotensinogen dan leptin yang berhubungan dengan peningkatan
insulin melalui GLUT 4, dan otot menyumbang sekitar 60-70% dari resistensi insulin, dan adiponektin dengan penurunan resistensi
penyerapan yang dimediasi insulin seluruh tubuh.34 Dalam keadaan insulin. TNFÿ dan IL-6 mengganggu pensinyalan insulin, lipolisis dan
makan, insulin meningkatkan sintesis glikogen melalui aktivasi fungsi endotel . . Produksi IL-6 ditingkatkan oleh aktivasi sistem saraf
glikogen sintase. Hal ini memungkinkan energi dilepaskan secara simpatis, misalnya stres.39 Depot jaringan adiposa berbeda dalam
anaerob melalui glikolisis, misalnya selama aktivitas otot yang intens. responsnya terhadap insulin.35 Adiposit dari penderita diabetes dan
Sel otot tidak bergantung pada glukosa (atau glikogen) untuk energi resisten insulin telah mengurangi translokasi GLUT 4, gangguan
selama keadaan basal, saat kadar insulin rendah. Insulin menekan pensinyalan intraseluler melalui penurunan gen IRS-1 dan ekspresi
katabolisme protein sementara defisiensi insulin mendorongnya, protein, gangguan PIP-3 kinase dan Akt (protein kinase B) yang
melepaskan asam amino untuk glukoneogenesis. Dalam kelaparan, distimulasi insulin.34
sintesis protein berkurang sebesar 50%.35 Meskipun data tentang
efek anabolik langsung dari insulin tidak konsisten, namun jelas
permisif, memodulasi status fosforilasi intermediet dalam jalur sintetik Hati
protein. Dalam studi eksperimental, dosis insulin yang mempromosikan Sementara penyerapan glukosa ke dalam hati tidak bergantung pada
sintesis protein secara signifikan lebih besar daripada dosis yang insulin, itu menyumbang sekitar 30% dari pembuangan glukosa yang
dibutuhkan untuk menekan proteolisis. Insulin bersifat anabolik dalam dimediasi insulin seluruh tubuh, 34 dengan insulin yang dibutuhkan
hubungannya dengan hormon pertumbuhan, IGF-1 dan asam amino untuk memfasilitasi proses metabolisme utama. Melalui pensinyalan
yang cukup.35 Pada resistensi insulin, sintesis glikogen otot terganggu; intraseluler yang dijelaskan di atas, sintesis glikogen distimulasi
ini tampaknya sebagian besar dimediasi oleh translokasi glukosa sementara sintesis protein dan metabolisme lipoprotein dimodulasi.30
intraseluler yang berkurang. Berkaitan dengan pergantian protein, Glukoneogenesis dan produksi badan keton dihambat.
satu penelitian melaporkan tidak ada perbedaan antara penderita Efek mitogenik insulin (dan hormon pertumbuhan) dimediasi melalui
diabetes tipe 2 yang resistan terhadap insulin dan kontrol, meskipun produksi faktor pertumbuhan seperti insulin di hati dan berpotensi
ini mengorbankan hiperinsulinemia dalam studi klem euglikemik melalui penekanan produksi sex-hormone binding globulin (SHBG).28
hiperinsulinemia ini.36
Jaringan Adiposa Meskipun pada defisiensi insulin, misalnya kelaparan, proses ini
Transpor glukosa intraseluler ke adiposit dalam keadaan postprandial dipengaruhi secara lebih seragam, hal ini belum tentu terjadi pada
bergantung pada insulin melalui GLUT 4; diperkirakan bahwa jaringan resistensi insulin. Hiperinsulinemia kompensasi, resistensi insulin
adiposa menyumbang sekitar 10% insulin yang merangsang diferensial, dan kebutuhan jaringan diferensial dapat memisahkan
pengambilan glukosa seluruh tubuh.34 Insulin merangsang proses ini.3 Resistensi terhadap
Klinik Biochem Rev Vol 26 Mei 2005 I 27
Machine Translated by Google
WilcoxG
efek metabolisme insulin menghasilkan peningkatan keluaran glukosa Hiperinsulinemia kompensasi yang menyertai resistensi insulin dikaitkan
melalui peningkatan glukoneogenesis (seperti pada kelaparan), namun, dengan peningkatan kadar faktor koagulan pro seperti PAI-1. Faktor-
tidak seperti kelaparan, hiperinsulinemia kompensasi menekan produksi faktor ini dianggap berkontribusi pada peningkatan agregasi trombosit
SHBG dan meningkatkan efek mitogenik insulin. yang terlihat pada keadaan resisten insulin. Sekresi endotelin 1 distimulasi
Perubahan dalam metabolisme lipoprotein merupakan manifestasi hati oleh insulin dan meningkat pada keadaan resisten insulin. Endotelin 1,
utama dari resistensi insulin. Peningkatan pengiriman asam lemak bebas, suatu vasokonstriktor kuat juga menghambat pensinyalan insulin melalui
dan penurunan katabolisme VLDL oleh adiposit yang resisten terhadap PIP-3 kinase dan bersaing dengan NO yang mengakibatkan disfungsi
insulin, menghasilkan peningkatan kandungan trigliserida hati dan sekresi endotel.
VLDL.38 Sintesis protein C reaktif hati, fibrinogen dan PAI-1 diinduksi Meskipun efek metabolik insulin dipengaruhi secara bervariasi pada
sebagai respons terhadap pro-infl turunan adiposit sitokin ammatory seperti resistensi insulin, sifat mitogenik, yang dimediasi melalui jalur kinase MAP
TNFÿ dan IL-6. Insulin juga dapat meningkatkan ekspresi gen faktor VII.39 (mitogen diaktifkan protein), tetap utuh. Efek mitogenik insulin pada
proliferasi sel otot polos endotel mungkin berkontribusi terhadap
aterosklerosis.33
Endotelium dan Pembuluh Darah
Insulin dan aksinya memainkan peran penting dalam berbagai aspek Otak
fungsi endotel, misalnya produksi oksida nitrat, sedangkan resistensi Sementara otak tidak tergantung pada insulin sejauh menyangkut
insulin sangat terkait dengan disfungsi endotel. Apakah asosiasi ini kausal, penyerapan glukosa intraseluler, reseptor insulin telah terletak di otak dan
atau dimediasi oleh mekanisme umum, masih menunggu klarifikasi. Fungsi terkonsentrasi di bola penciuman, hipotalamus, hippocampus, 40 retina
sel endotel vaskular sangat penting untuk banyak aspek biologi dan pembuluh pleksus koroid, serta di daerah striatum dan korteks serebral
kardiovaskular, dengan disfungsi endotel terlihat pada tahap awal misalnya lobus temporal medial. Insulin diyakini bertindak sebagai
aterosklerosis dan faktor risiko klinis yang terkait. Sel endotel tidak hanya neuropeptida, terlibat dalam rasa kenyang, pengaturan nafsu makan,
menyediakan lapisan fisik pembuluh darah tetapi juga mengeluarkan penciuman, memori dan kognisi.41 Insulin dapat diangkut secara aktif dari
berbagai faktor yang mempengaruhi tonus pembuluh darah, fungsi aliran darah atau disintesis secara lokal.
trombosit, koagulasi dan fibrinolisis. Masalah klinis berkembang ketika
proses ini tidak seimbang. Insulin juga tampaknya bertindak melalui neurotransmiter dan peptida
pengatur nafsu makan lainnya. Leptin dan insulin tampaknya memiliki jalur
pensinyalan yang sama di hipotalamus.39 Ada dugaan adanya hubungan
potensial dengan penyakit Alzheimer, mengingat peran insulin dalam
Nitrit oksida (NO) adalah faktor utama dalam arteri besar yang memediasi fungsi kognitif normal dan dalam pengaturan protein prekursor amiloid dan
relaksasi yang bergantung pada endotel. Ini juga menghambat agregasi beta-amiloid itu sendiri.42 Kemungkinan bahwa sindrom yang terkait
trombosit, adhesi sel dan proliferasi sel otot polos. dengan resistensi insulin di perifer, seperti obesitas dan diabetes tipe 2,
NO disintesis dari L-arginine, molekul oksigen dan NADPH, melalui aktivitas juga dapat dikaitkan dengan resistensi insulin di otak, dengan disregulasi
enzim endotel nitric oxide synthase (eNOS), dan kofaktornya nafsu makan dan berat badan, sangat menarik.41
tetrahydrobiopterin, flavin adenine dinucleotide dan flavin mononucleotide.
Menariknya, arginin adalah secretatogue kuat untuk insulin dan ada jalur
umum terakhir untuk pensinyalan intraseluler dari eNOS dan insulin. Insulin
meningkatkan produksi tetrahydrobiopterin dengan menstimulasi enzim Pankreas
biosintetik GTP cyclohydrolase, dan menstimulasi eNOS dengan fosforilasi Sel ÿ pankreas memiliki reseptor untuk insulin dan IGF 1. Insulin mungkin
eNOS yang tidak bergantung kalsium pada residu serin dan treonin melalui memiliki peran dalam mengatur sekresi insulin yang distimulasi oleh
PIP-3 kinase dan Akt glukosa, kemungkinan melalui sensor glukosa atau pertumbuhan sel ÿ.43
(protein kinase B). Dengan demikian produksi oksida nitrat ditingkatkan.
Insulin juga mendorong pelepasan endotelin vasokonstriktor sementara Reseptor
TNFÿ menurunkan ekspresi eNOS dan menginduksi pelepasan Faktor von insulin hipofisis ditemukan di kelenjar hipofisis anterior44 dan ditemukan
Willebrand. Pada resistensi insulin, kadar tetrahidrobiopterin berkurang, berhubungan dengan ÿ endorphin, konsisten dengan peran dalam
jalur stimulasi eNOS diatur turun, dan respons vasodilator terhadap insulin pengaturan asupan makanan.45 Insulin bekerja bersamaan dengan
dan agonis kolinergik terganggu. Kemampuan insulin untuk menetralkan hormon lainnya. Insulin merangsang produksi hormon pertumbuhan dari
defosforilasi Akt yang dimediasi TNFÿ dalam sel endotel juga hilang. kelenjar hipofisis, yang pada gilirannya mendorong produksi IGF-1 oleh
hati.
Asam lemak bebas, meningkat pada keadaan resisten insulin, juga
menghambat aktivitas eNOS, menurunkan produksi NO.
28 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
Ginjal Diet tinggi karbohidrat umumnya dikaitkan dengan peningkatan
Ginjal tidak memerlukan insulin untuk transpor glukosa. sensitivitas insulin dalam jangka pendek. Sekresi insulin tergantung
Reseptor insulin ditemukan di tubulus proksimal; insulin mengatur pada jenis dan bentuk fisik dari karbohidrat yang dikonsumsi.
transportasi mineral dan glukoneogenesis di ginjal. Indeks glikemik (GI) suatu makanan mengacu pada area di bawah
Insulin terdegradasi di ginjal.46 In vivo, konsentrasi fisiologis insulin kurva untuk konsentrasi glukosa darah versus waktu, relatif terhadap
mengurangi ekskresi natrium urin.47 dosis glukosa yang setara. GI lebih rendah di mana pencernaan,
penyerapan dan/atau konversi menjadi glukosa terjadi lebih lambat.
Gonad Tingkat sekresi insulin cenderung lebih rendah ketika GI rendah
Reseptor insulin ditemukan di ovarium dan tampaknya memiliki peran makanan dikonsumsi. Tidak semua karbohidrat (dalam bentuk yang
dalam meningkatkan steroidogenesis.48 Insulin dan IGF-1 bekerja dimurnikan) setara dalam respons insulinnya.58 Sebagai contoh,
secara sinergis dengan FSH untuk meningkatkan produksi estrogen fruktosa cenderung tidak menimbulkan respons insulin yang nyata
oleh sel granulosa dan dengan LH untuk meningkatkan produksi seperti glukosa. Overfeeding kronis dengan sukrosa telah dilaporkan
androgen oleh sel stroma tekak.49 Mereka juga memiliki telah meningkatkan deposisi jaringan adiposa visceral pada hewan pengerat,
ditemukan pada testis tikus dimana insulin dapat bekerja secara yang mungkin memiliki implikasi jangka panjang untuk resistensi
sinergis dengan LH pada steroidogenesis.50 insulin.59 Sementara ada kekhawatiran bahwa diet karbohidrat tinggi
dapat memperburuk manifestasi klinis dari resistensi insulin.
Tulang sindrom, hal ini mungkin tergantung pada jenis karbohidrat dan asupan
Insulin tampaknya anabolik dalam tulang. Reseptor insulin ditemukan serat makanan yang memiliki efek menguntungkan.60
di osteoblas dan osteoklas. Insulin merangsang tulang
pembentukan oleh osteoblas dan juga dilaporkan menekan fungsi Serat makanan memiliki efek tidak langsung pada sekresi dan kerja
osteoklas.51 insulin. Efek pada motilitas usus dan waktu transit, sekresi hormon
gastrointestinal misalnya GIP,61 GLP-1,62 dan fermentasi kolon yang
Pengaruh Fisiologis pada Aksi Insulin dan Insulin menghasilkan asam lemak rantai pendek yang menekan
Perlawanan glukoneogenesis hepatik, semuanya telah dilaporkan.63 Pati resisten,
Diet dan Kelaparan yang tidak dicerna di usus halus tetapi lolos ke usus besar bertindak
Kadar insulin rendah pada keadaan basal, seperti selama kelaparan, sebagai substrat untuk fermentasi kolon (seperti serat makanan) dan
memfasilitasi mobilisasi asam lemak dan gliserol dari jaringan adiposa telah dilaporkan menurunkan glukosa post prandial dan respons
dan asam amino dari otot.29 Sementara kadar insulin biasanya lebih insulin, meningkatkan sensitivitas insulin seluruh tubuh, menurunkan
tinggi pada keadaan kenyang, derajat sensitivitas insulin dapat trigliserida dan HDL, meningkatkan rasa kenyang dan mengurangi
dipengaruhi oleh komposisi dietnya. Konsumsi energi berlebih yang penyimpanan lemak.64
kronis meningkatkan hiperinsulinemia dan resistensi insulin melalui
stimulasi sekresi insulin, sintesis trigliserida, dan akumulasi lemak Konsumsi protein merangsang sekresi insulin, mendukung anabolisme;
dengan regulasi penurunan reseptor insulin dan pensinyalan pasca itu mungkin memediasi efeknya pada rasa kenyang dengan cara ini.
reseptor.17 Ini juga merangsang sekresi glukagon dari sel ÿ pankreas dan
mendorong glukoneogenesis.65
Diet tinggi lemak cenderung dikaitkan dengan resistensi insulin, Konsumsi alkohol moderat telah dilaporkan meningkatkan peningkatan
khususnya yang berkaitan dengan lemak jenuh52 dan asam lemak insulin postprandial dan mengurangi peningkatan glukosa postprandial
trans.53 Komposisi asam lemak dianggap berperan dalam setelah makan rendah karbohidrat.66 Studi populasi melaporkan kurva
perkembangan resistensi insulin jangka panjang, melalui efek pada berbentuk U sehubungan dengan tingkat konsumsi alkohol dan
komposisi lipid membran. . Asam lemak tak jenuh ganda rantai panjang resistensi insulin.67
memainkan peran fisiologis dalam menjaga fluiditas membran sel dan
pensinyalan sel; mereka juga memengaruhi ekspresi gen dan Mikronutrien yang berpotensi penting termasuk seng, kromium, dan
merupakan ligan endogen untuk reseptor proliferator peroksisom.54 mungkin besi. Seng tampaknya penting dalam biosintesis dan sekresi
Studi epidemiologis melaporkan hubungan antara rasio asam lemak insulin, dan terkonsentrasi di pankreas. Defisiensi kromium telah
omega 6 dengan omega 3 dan prevalensi diabetes tipe 2.55 Ada dikaitkan dengan intoleransi glukosa dan resistensi insulin pada pasien
beberapa penelitian eksperimental untuk mendukung hipotesis ini; dengan nutrisi parenteral jangka panjang, meskipun pekerjaan di
meskipun asam lemak omega-3 rantai panjang dalam dosis bidang ini terhambat oleh kesulitan dalam pengukuran yang akurat
farmakologis dianggap mengganggu toleransi glukosa,56 meta-analisis dari konsentrasi jejak elemen ini dalam cairan tubuh.68 Bentuk aktif
tidak mengkonfirmasi hal ini.57 kromium adalah oligopeptida yang disebut chromomodulin yang
meningkatkan aktivitas tirosin kinase dari
Klinik Biochem Rev Vol 26 Mei 2005 I 29
Machine Translated by Google
WilcoxG
reseptor insulin. Ion kromium diangkut dalam aliran darah pada stres psikososial dalam perkembangan sindrom metabolik baru-baru
transferin.69 Akumulasi besi dikaitkan dengan gangguan sensitivitas ini ditinjau.83
insulin dan diabetes tipe 2, dan ciri-ciri sindrom metabolik.70 Apakah
efek merugikan dari akumulasi besi diperparah oleh potensi efek Tidur dan Kurang Tidur
merusak yang dihasilkan pada kromium metabolisme memiliki Kurang tidur akut pada orang dewasa muda yang sehat telah
dilaporkan meningkatkan konsentrasi glukosa darah puasa dalam
belum ditangani. Selain efek nutrisi hubungannya dengan perubahan sekresi kortisol diurnal dan berkurangnya
komponen diet, makanan individu dan fitokimia komponennya juga variabilitas denyut jantung.84 Efek ini menunjukkan peningkatan
dapat mempengaruhi sekresi dan kerja insulin.71-73. sekresi hormon counter-regulatory melalui hyper-arousal dengan
aktivasi aksis adrenal hipotalamus-hipofisis.85 Ada juga kumpulan
bukti bahwa kurang tidur kronis dapat berdampak pada insulin dan
Latihan & Aktivitas Fisik resistensi insulin. Studi epidemiologi baru-baru ini melaporkan bahwa
Sejak pengamatan luar biasa Chauveu & Kaufman di berkurangnya durasi tidur dikaitkan dengan peningkatan BMI.86
1887 bahwa “Ketika seekor kuda mengunyah jerami, konsentrasi Kurang tidur dikaitkan dengan penurunan konsentrasi plasma leptin,
glukosa dalam darah yang menguras otot masseternya secara hormon peptida adiposit yang mengatur massa lemak dan nafsu
substansial menurun” 74 banyak bukti mendukung peran olahraga makan, dan peningkatan konsentrasi ghrelin, yang meningkatkan
dalam meningkatkan sensitivitas insulin dan hasil yang bermanfaat nafsu makan.87 Hormon pertumbuhan adalah disekresikan selama
dalam keadaan resisten insulin. Studi epidemiologis seperti US tidur gelombang lambat, tidur menurun dengan bertambahnya usia88
Physicians Health Study telah melaporkan penurunan substansial dan defisiensi hormon pertumbuhan pada orang dewasa telah
dalam risiko relatif diabetes tipe 2 dengan aktivitas fisik teratur dikaitkan dengan adipositas sentral dan resistensi insulin,89 tetapi
seumur hidup.75 Uji klinis terkontrol acak berskala besar seperti apakah kurang tidur bekerja melalui mekanisme ini tidak ditetapkan
Diabetes Prevention dengan jelas.
Program76 dan Studi Pencegahan Finlandia77 menunjukkan Apnea tidur obstruktif (OSA), di mana gangguan tidur diakibatkan
penurunan 58% dalam perkembangan gangguan toleransi glukosa oleh obstruksi pernapasan selama tidur, dikaitkan dengan gangguan
menjadi diabetes tipe 2 dengan modifikasi gaya hidup intensif yang toleransi glukosa terlepas dari adipositas,90 dan membaik dengan
mencakup minimal 20-30 menit olahraga per hari. perawatan tekanan jalan napas positif terus menerus91 tetapi apakah
Latihan akut meningkatkan translokasi GLUT 4 menjadi sarcolemmal ini karena resolusi hipoksia dan hiperkapnia, atau untuk efek pada
membran, sedangkan latihan olahraga kronis meningkatkan ekspresi kualitas tidur, tidak jelas.
mRNA Glut 4.78 Selain mekanisme yang bergantung pada insulin ini,
peningkatan penyerapan glukosa ke dalam otot yang berolahraga Kehamilan
terjadi melalui mekanisme independen insulin ganda.79 Latihan Kehamilan normal ditandai dengan resistensi insulin yang terbesar
olahraga tampaknya meningkatkan sensitivitas insulin dengan pada trimester ketiga. Ini tampaknya merupakan respons adaptif,
meningkatkan pensinyalan insulin pasca-reseptor; 80 peningkatan mengalihkan glukosa dan lipid ke janin yang sedang berkembang92
transportasi glukosa yang dimediasi insulin tampaknya terkait dengan dan berpikir karena efek gabungan dari laktogen plasenta manusia,
peningkatan transduksi sinyal pada tingkat protein IRS dan PI 3-kinase.79 progesteron, estradiol dan kortisol, yang bertindak sebagai hormon
kontra-regulasi terhadap insulin.
Berlebihan dari resistensi insulin biasanya terlihat pada kehamilan
menekankan
dikaitkan dengan diabetes mellitus gestasional dan hipertensi
Resistensi insulin biasanya terlihat pada stres katabolik penyakit gestasional.93
parah dengan implikasi morbiditas dan mortalitas.
Mekanisme termasuk aktivasi aksis hipotalamo-hipofisis adrenal Obesitas
(HPA) yang menghasilkan peningkatan nyata hormon pengatur, Peningkatan jaringan adiposa, terutama di tubuh bagian atas atau
serta efek sitokin inflamasi.81 Yang terakhir merusak pensinyalan deposisi “android”, pertama kali dikaitkan dengan diabetes dan
reseptor insulin di otot rangka, hati, dan jaringan adiposa; zat antara penyakit pembuluh darah oleh ahli endokrin Perancis Jean Vague
seperti IkB kinase, MAP kinase, ceramide, dan ganglioside GM3 pada tahun 1956.94 Resistensi insulin meningkat dengan
memengaruhi beberapa jalur transduksi sinyal utama. Contoh penting meningkatnya indeks massa tubuh, lingkar pinggang dan khususnya
termasuk penghambatan aktivitas reseptor insulin kinase melalui rasio pinggang- pinggul 0,95 Ini mencerminkan peningkatan adipositas
fosforilasi serin IRS-1, dengan efek hilir pada PI-3 kinase, dan dengan terutama peningkatan kadar jaringan adiposa visceral. Jaringan
demikian translokasi GLUT 4, serta efek langsung pada PI-3 kinase, adiposa visceral mengacu pada lemak intra perut di sekitar usus dan berkorelasi d
Akt/protein kinase B dan protein kinase C.82 Signifikansi potensi gemuk. Jaringan adiposa visceral memiliki karakteristik metabolik
kronis yang berbeda dengan lemak subkutan. Ini lebih aktif secara metabolik
sehubungan dengan pergantian asam lemak bebas; yang meningkat
30 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
fluks asam lemak bebas meningkatkan resistensi insulin pada tingkat sel 2 diabetes di mana sisa fungsi sel ÿ ada. Agen lain bertindak dengan
dan meningkatkan produksi VLDL hati.35 Pre adiposit omental telah mengurangi resistensi insulin. Metformin, sebuah biguanida, mengurangi
meningkatkan aktivitas 11-ÿ hidroksisteroid dehidrogenase tipe 1, awalnya keluaran glukosa hati dan pada tingkat yang lebih rendah resistensi insulin
diduga meningkatkan resistensi insulin melalui efek glukokortikoid aktif yang perifer dan digunakan pada diabetes tipe 2 dan sindrom ovarium polikistik
diproduksi secara lokal, melalui konversi kortison menjadi kortisol, meskipun (PCOS). Tindakan selulernya kurang dicirikan dengan baik. Baru-baru ini,
signifikansi temuan in vivo ini masih diperdebatkan.96 kelas agonis untuk reseptor gamma proliferator peroksisom telah tersedia
dan agen ini lebih kuat daripada metformin dalam mengurangi resistensi
insulin perifer. Mereka mempromosikan diferensiasi adiposit subkutan dan
Jaringan adiposa menghasilkan sejumlah sitokin yang dikaitkan dengan mendukung redistribusi trigliserida dari depot lemak adiposa hati dan visceral
resistensi insulin, termasuk sitokin dengan aktivitas pro-inflamasi misalnya ke perifer. Mereka meningkatkan resistensi insulin di jaringan adiposa tetapi
TNFÿ, interleukin, dan PAI-1. Ada perbedaan regional dalam produksi sitokin tidak di otot. Thiazolidinediones seperti rosiglitazone dan pioglitazone
adiposit. Menariknya laporan baru-baru ini menunjukkan peningkatan digunakan pada diabetes tipe 2103 dan telah efektif pada PCOS.28 Namun,
resistensi insulin dengan aspirin dosis tinggi, mendukung hipotesis penulis ada kekhawatiran mengenai keamanan thiazolidinediones pada kehamilan;
mengenai peran serine kinase IkB kinase-ÿ (IKK-Eÿ) sebagai mediator apalagi hasil kehamilan jangka panjang dari semua agen sensitisasi insulin
resistensi insulin intraseluler pada otot. Resistensi insulin otot terkait ini, termasuk metformin, tidak diketahui.104
dengan peningkatan trigliserida intramyoseluler, berasal dari lipolisis jaringan
adiposa.97.
Sindrom Resistensi Insulin Sindrom
Resistensi insulin yang terlihat pada obesitas diyakini terutama melibatkan resistensi insulin menjelaskan sekelompok kelainan yang lebih sering terjadi
otot dan hati, dengan peningkatan asam lemak bebas turunan adiposit yang pada individu yang resisten insulin. Ini termasuk intoleransi glukosa,
mendorong akumulasi trigliserida dalam jaringan ini.97 Hal ini lebih mungkin dislipidemia, disfungsi endotel dan peningkatan faktor prokoagulan,
terjadi pada adiposit yang resisten terhadap insulin.32 Fluks asam lemak perubahan hemodinamik, peningkatan penanda inflamasi, metabolisme
bebas lebih besar dari visceral jaringan adiposa dan lebih mungkin pada asam urat abnormal, peningkatan sekresi testosteron ovarium dan gangguan
orang-orang dengan resistensi insulin adiposit yang dimediasi secara pernapasan saat tidur. Sindrom klinis yang terkait dengan resistensi insulin
genetik.98 Meskipun perbedaan individu dalam efek peningkatan adipositas termasuk diabetes tipe 2, penyakit kardiovaskular , hipertensi esensial,
ada, penambahan berat badan memburuk dan penurunan berat badan sindrom ovarium polikistik, penyakit hati berlemak non-alkohol, beberapa
meningkatkan resistensi insulin pada mereka yang memiliki kecenderungan demikian.99
bentuk kanker, dan sleep apnea.3
Pengaruh farmakologis terhadap Tindakan Insulin dan Insulin
Perlawanan
Berbagai agen farmakologis telah dikaitkan dengan gangguan toleransi Sindrom Klinis Terkait dengan Resistensi Insulin Diabetes tipe 2 dan
glukosa. Agen antihipertensi seperti diuretik dan ÿ-blocker, kortikosteroid, Sindrom Metabolik akan menjadi sindrom klinis yang paling umum terkait
kontrasepsi oral, asam nikotinat dan agen antipsikotik telah dilaporkan dengan resistensi insulin. Lainnya termasuk hipertensi, PCOS, penyakit hati
mengganggu toleransi glukosa100,101 seperti halnya anti-retroviral protease berlemak non-alkohol, beberapa bentuk kanker dan OSA,3 yang oleh
inhibitor yang digunakan untuk mengobati infeksi human immunodeficiency beberapa penulis dianggap sebagai komponen dari sindrom metabolik itu
virus.102 Mekanismenya bervariasi ; ÿ-blocker merusak sekresi insulin dari sendiri. Ada juga kondisi yang relatif umum di mana resistensi insulin
pankreas dengan blokade ÿ-adrenoceptors, diuretik thiazide dianggap merupakan fenomena sekunder; ini termasuk penyakit akut, sirosis hati,
bekerja dengan mengurangi kadar kalium, kortikosteroid dan kontrasepsi gagal ginjal, kehamilan, hipertiroidisme, penyakit Cushing dan sindrom
oral memiliki aktivitas hormonal kontra-regulasi,101 dan inhibitor protease Cushing serta acromegaly dan phaeochromocytoma yang kurang umum.26
HIV-1 menghasilkan lipodistrofi parsial . dengan hilangnya lemak subkutan Dalam banyak dari ini, resistensi insulin disebabkan peningkatan produksi
perifer dan akumulasi jaringan adiposa tubuh yang menyebabkan resistensi hormon counter-regulatory . .
insulin.102
Namun ada juga sejumlah besar kelainan yang jarang terjadi di mana
Selain insulin itu sendiri, sejumlah agen digunakan secara terapeutik untuk resistensi insulin merupakan gambaran klinis utama.28,105 (Tabel 3)
memodulasi kerja insulin. Ini termasuk sulfonilurea, yang menstimulasi Meskipun jarang terjadi secara individual, kondisi ini dapat memberikan
sekresi insulin dari sel ÿ melalui aktivasi reseptor sulfonilurea yang wawasan tentang mekanisme resistensi insulin di tempat lain. Biasanya
berdampingan dengan saluran K+ -ATP pada membran sel.12 Mereka mereka ditandai dengan gangguan pada sistem organ dimana aksi insulin
digunakan dalam tipe memainkan peran penting.
Klinik Biochem Rev Vol 26 Mei 2005 I 31
Machine Translated by Google
WilcoxG
Kondisi Jarang Terkait dengan Resistensi Insulin Kondisi Umum Terkait dengan Resistensi Insulin
Lipodistrofi mewakili sekelompok kelainan yang ditandai dengan Diabetes Tipe 2 (T2DM)
kurangnya jaringan adiposa subkutan, yang dapat diwariskan atau Mengikuti karya perintis oleh Bornstein112 dan karya pemenang
didapat (misalnya sekunder akibat pengobatan dengan penghambat Hadiah Nobel dari Yalow dan Berson,113 tes insulin pertama menjadi
protease untuk HIV). Mereka terkait dengan akumulasi lemak di hati tersedia secara luas pada akhir 1960-an;28 kemudian dikonfirmasi
dan otot dan resistensi insulin. Pencangkokan simpanan jaringan bahwa pasien diabetes dengan apa yang disebut atau onset maturitas
adiposa subkutan telah terbukti meningkatkan resistensi insulin pada atau diabetes tipe 2 memiliki kadar insulin plasma normal atau
model hewan dari gangguan ini. Untuk meningkat. Resistensi insulin dilaporkan menjadi ciri khas T2DM pada
ulasan terbaru tentang topik ini lihat Garg & Misra.106 awal tahun 1970-an.3 Ketidakmampuan progresif sel ÿ untuk
mengkompensasi resistensi insulin yang ada dengan hiperinsulinemia
Gangguan metabolisme yang diturunkan seperti gangguan mitokondria yang cukup, menandakan onset klinis dari gangguan ini.3 Sementara
dan penyakit penyimpanan glikogen mempengaruhi otot dan hati pada studi kembar
khususnya, dan masing-masing berhubungan dengan diabetes dan dan analisis keterkaitan konsisten dengan komponen genetik yang
resistensi insulin. Pada gangguan mitokondria sering terjadi kegagalan kuat dalam perkembangan diabetes tipe 2, beberapa dekade penelitian
pankreas yang mencerminkan kegagalan pembentukan energi sel ÿ gagal mengidentifikasi kelainan genetik yang dominan pada sebagian
yang berkontribusi terhadap perkembangan diabetes; 107 namun besar kasus.26 Etiologi T2DM dianggap poligenik, dengan faktor
gangguan oksidasi asam lemak mungkin lingkungan ditumpangkan pada kecenderungan dasar ini.
berkontribusi terhadap defek pada aksi insulin pada tingkat seluler.97
Menariknya, pasien diabetes dengan MELAS (Myoclonus, Elipepsy,
Lactic Acidosis, dan Stroke-like episode) terlihat lebih baik dengan diet Resistensi insulin biasanya mendahului perkembangan diabetes dan
tinggi lemak,108 tetapi juga diet rendah karbohidrat, yang meminimalkan umumnya ditemukan pada kerabat tingkat pertama yang tidak
akumulasi laktat akibat metabolisme piruvat yang tidak sempurna di terpengaruh.28 Morbiditas gangguan ini berhubungan dengan tingkat
Siklus Kreb. Baru-baru ini peningkatan lipid intramioseluler dan keparahan hiperglikemia dan konsekuensi metabolik dari resistensi
penurunan fosforilasi mitokondria telah dilaporkan pada keturunan insulin itu sendiri. Defek utama pada aksi insulin tampak pada sel otot
pasien dengan diabetes tipe 2 yang resistan terhadap insulin, dan adiposit, dengan gangguan translokasi GLUT 4 yang menyebabkan
menunjukkan disfungsi mitokondria halus mungkin lebih luas pada gangguan transpor glukosa yang diperantarai insulin.28
gangguan ini. Pada penyakit penyimpanan glikogen tipe I, manifestasi
dewasa mencolok secara klinis untuk tumpang tindih mereka dengan
Sindrom Metabolik.105 Resistensi insulin dalam kelompok ini baru-baru Hiperinsulinemia kompensasi berkembang pada awalnya, tetapi fase
ini telah didokumentasikan,110 dan kemungkinan berhubungan dengan pertama sekresi insulin hilang pada awal gangguan.
akumulasi lemak hati yang terjadi pada kondisi ini. Stres lingkungan dan fisiologis tambahan seperti kehamilan,
penambahan berat badan, aktivitas fisik dan obat-obatan dapat
Topik sindrom genetik yang menyebabkan diabetes, termasuk yang memperburuk resistensi insulin. Karena sel ÿ gagal mengkompensasi
menyebabkan resistensi insulin, telah ditinjau secara ekstensif oleh resistensi insulin yang ada, gangguan toleransi glukosa dan diabetes
Scheuner et al.111 berkembang. Saat kadar glukosa meningkat, fungsi sel ÿ semakin
memburuk, dengan penurunan sensitivitas terhadap glukosa dan
hiperglikemia yang memburuk. Massa sel pulau pankreas dilaporkan
Tabel 3. Kelainan genetik yang jarang terkait dengan resistensi
berkurang ukurannya pada pasien diabetes; faktor humoral dan
insulin.26,29
endokrin mungkin penting dalam mempertahankan massa sel islet.11
Berbeda dengan sebagian besar bentuk diabetes tipe 2, dasar genetik
Sindrom Down
Sindrom Turner dari Maturity Onset diabetes of the Young (MODY) telah dicirikan
Sindrom Klinefelter dengan baik dan berhubungan dengan defek pada glukokinase, maka
Thalassemia intraseluler transpor glukosa.26
Haemochromatosis
Lipodystrophy
Progeria Sindrom Metabolik
Chorea Huntington Pengamatan awal Jean Vague pada tahun 1950-an ditemukan kembali
Myotonic dystrophy sekitar tiga dekade kemudian, ketika pada tahun 1988 diusulkan bahwa
Ataksia Friedrich
individu dengan intoleransi glukosa, peningkatan trigliserida, kolesterol
Sindrom Laurence-Moon-Biedel
HDL rendah, dan hipertensi esensial berada pada risiko kardiovaskular
Penyakit penyimpanan glikogen tipe I & III
yang sangat meningkat. Hubungan yang mendasari dengan insulin
Gangguan mitokondria (?)
perlawanan tidak sepenuhnya dihargai
32 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
pada saat itu dan awalnya disebut sebagai Sindrom X, tetapi Hipertensi
kemudian sebagai Sindrom Metabolik. Panel Perawatan Dewasa Hipertensi esensial telah dikaitkan dengan resistensi insulin
III dari Panel Penasihat Kolesterol Nasional mendefinisikan hingga 50% kasus.117 Terdapat korelasi yang kuat antara
sindrom metabolik sebagai "konstelasi faktor risiko lipid dan non tekanan darah dengan berat badan. Signifikansi resistensi insulin
lipid yang berasal dari metabolisme". Kriteria diagnostik mereka pada hipertensi masih menjadi kontroversi, karena 50% hipertensi
untuk sindrom metabolik memerlukan tiga hal berikut: esensial tampaknya tidak memiliki resistensi insulin. Mekanisme
yang diusulkan termasuk peningkatan retensi natrium ginjal dan
Obesitas perut peningkatan aktivitas sistem saraf simpatik dari hiperinsulinemia
Pria: lingkar pinggang >40 inci (102 cm) kompensasi. Namun disfungsi endotel dari resistensi terhadap
Wanita: lingkar pinggang >35 inci (88 cm) pembentukan oksida nitrat yang dimediasi insulin dianggap
Glukosa puasa ÿ 110 <126 mg/dL (6.1-7.0 mmol/L) memiliki signifikansi yang lebih jelas.118,119
Tekanan darah ÿ 130/80 mmHg
Trigliserida >150 mg/dL (>1,7 mmol/L)
kolesterol HDL PCOS
Pria <40 mg/dL (<1,0 mmol/L) Bukti yang meyakinkan menghubungkan resistensi insulin dengan
Wanita <50 mg/dL (<1,3 mmol/L) patogenesis ini, kelainan endokrin yang paling umum pada wanita
premenopause.3 Keparahan defek genetik pada sensitivitas
Diperkirakan bahwa di Amerika Serikat sekitar 25% insulin mungkin dapat menentukan kemungkinan berkembangnya
orang dewasa memiliki sindrom.3 Selain predisposisi genetik PCOS untuk indeks massa tubuh tertentu. Defek transduksi
yang mendasari resistensi insulin, aktivitas fisik dan kelebihan sinyal insulin yang ditentukan secara genetik telah dijelaskan
asupan energi dianggap sebagai faktor risiko utama, dengan pada PCOS, dengan resistensi insulin mendahului manifestasi
kontribusi yang tidak diketahui dari faktor lain seperti klinis. Kerusakan lebih lanjut pada insulin
stres kronis114 dan kurang tidur.115 sensitivitas terjadi dengan percepatan pertumbuhan remaja dan
perkembangan obesitas.120 Disfungsi ovarium berkaitan dengan
Dislipidemia efek kompensasi hiperinsulinemia yang meningkatkan sekresi LH
Kelainan lipid yang berhubungan dengan resistensi insulin hipofisis dan produksi androgen oleh sel-sel teka ovarium.
mempengaruhi semua fraksi lipid. Mereka dicirikan oleh Aromatisasi androgen dalam pengaturan obesitas meningkatkan
peningkatan kadar trigliserida puasa, peningkatan lipoprotein produksi estrogen, selanjutnya merusak fungsi aksis gonad
sisa kaya trigliserida postprandial, kolesterol HDL rendah, dan hipotalamo-hipofisis.
partikel LDL padat kecil. Pola ini berkorelasi kuat dengan risiko Hiperinsulinemia juga menekan produksi SHBG oleh hati,
kardiovaskular, dan pengobatan menurunkan risiko ini.3 selanjutnya meningkatkan androgen bebas. Peningkatan
Apolipoprotein B (apo B) biasanya dikaitkan dengan kolesterol androgen pada gilirannya semakin memperburuk resistensi
LDL; namun, pada dislipidemia aterogenik yang resistan terhadap insulin.120 Modalitas terapi yang mengurangi resistensi insulin
insulin, sebagai respons terhadap peningkatan pengiriman asam biasanya menghasilkan perbaikan fungsi ovarium. Ini termasuk
lemak bebas ke hati, terjadi kelebihan produksi apo B dan penurunan berat badan, olahraga, dan agen farmakologis yang
peningkatan sintesis trigliserida VLDL. Ini termasuk partikel VLDL peka terhadap insulin seperti metformin dan troglitazone.28
yang mengandung apo B. Kadar kolesterol HDL yang rendah
merupakan faktor risiko independen untuk penyakit kardiovaskular; Penyakit Hati Berlemak Nonalkohol (NAFLD)
HDL rendah yang terlihat pada dislipidemia aterogenik NAFLD semakin dikenal dengan berbagai negara yang
berhubungan dengan ukuran partikel HDL yang berkurang. melaporkan tingkat prevalensi 10-24% dari populasi umum;
Ukuran partikel HDL yang rendah berkaitan dengan pertukaran namun di antara orang gemuk, 50-75% terpengaruh. 25-50%
trigliserida VLDL untuk ester kolesterol dalam LDL dan HDL anak obesitas terkena dampaknya. NAFLD dapat berkembang
melalui protein transfer ester kolesterol. Ketika trigliserida dalam menjadi steatohepatosis non-alkohol, steatohepatitis, fibrosis
LDL dan HDL menjalani hidrolisis, LDL dan HDL terdeplesi atau sirosis, yang menunjukkan peningkatan kerusakan hati.
kolesterol kecil tetap ada.116 Partikel LDL padat kecil dilaporkan Selama masa tindak lanjut 3,5-11 tahun, 28% pasien dengan
lebih aterogenik mungkin karena kecenderungannya yang NAFLD mengalami kerusakan hati yang progresif.121 Resistensi
meningkat terhadap oksidasi dan proporsi apo B yang lebih insulin perifer pada lemak dan otot menyebabkan peningkatan
besar. Dibandingkan dengan non-insulin keadaan resisten, kadar pengiriman asam lemak bebas ke hati, meningkatkan sintesis
kolesterol LDL tertentu mewakili jumlah apo B yang lebih besar trigliserida. Perlemakan hati berkembang ketika sintesis trigliserida
yang mengandung partikel LDL padat kecil dan ini meningkatkan risiko.107
hati melebihi sintesis hati dan ekspor trigliserida VLDL.3
Klinik Biochem Rev Vol 26 Mei 2005 I 33
Machine Translated by Google
WilcoxG
Kanker metode termasuk penilaian resistensi insulin berdasarkan
Resistensi insulin dengan hiperinsulinemia kompensasi telah terlibat pemodelan matematika meliputi Homeostasis Assessment Model
dalam etiologi kanker tertentu, termasuk kanker usus besar, (HOMA)129 dan Quantitative Insulin Sensitivity Check Index
endometrium, mungkin sel pankreas dan ginjal122 dan kanker (QUICKI), Continuous Infusion of Glucose with Model Assessment
payudara.3 Dalam kasus kanker kolon diusulkan bahwa resistensi (CIGMA), Tes Toleransi Glukosa Intravena Sampel Sering dan
insulin menyebabkan hiperinsulinemia dengan peningkatan IGF. -1 pemodelan minimal.
tingkat mempromosikan proliferasi sel epitel usus; perubahan pada Metode-metode ini telah diulas baru-baru ini.130
NF-ÿB dan pensinyalan reseptor teraktivasi proliferator peroksisom
juga dapat memengaruhi kinetika kolonosit.123 Selain efek mitogenik Metode Klinis
insulin itu sendiri, efek resistensi insulin pada fungsi ovarium dan Sebagian besar metode yang diuraikan di atas memiliki penerapan
metabolisme hormon seks, bersama dengan tingkat SHBG yang yang terbatas dalam pengaturan klinis. Pendekatan yang paling
rendah berpotensi meningkatkan bio- tingkat hormon yang tersedia, banyak digunakan adalah mengukur insulin plasma puasa.
juga dapat berkontribusi terhadap karsinogenesis.124 Sementara informasi tambahan dapat diperoleh dengan HOMA dan
QUICKI, dengan memasukkan glukosa puasa simultan ke dalam
persamaan, telah diperdebatkan bahwa sebagian besar varian
OSA resistensi insulin tercermin dalam tingkat insulin saja.131 Belfi ore
OSA umumnya dianggap sebagai komplikasi obesitas yang et al telah mengembangkan Sensitivitas Insulin Indeks berdasarkan
dimanifestasikan oleh obstruksi mekanis pada saluran pernapasan insulin basal dan yang diinduksi OGTT, glukosa dan kadar asam
bagian atas saat tidur. Namun ada beberapa bukti yang menunjukkan lemak bebas; ini adalah ukuran praktis resistensi insulin seluruh
bahwa OSA mungkin merupakan kondisi sistemik yang berhubungan tubuh dalam pengaturan klinis dan unik dalam mengukur sensitivitas
dengan resistensi insulin.3 Namun fragmentasi tidur dan hyperarousal terhadap penekanan kadar asam lemak bebas yang dimediasi insulin.
yang dihasilkan dapat meningkatkan resistensi insulin melalui Mereka dapat diterapkan pada kadar glukosa dan insulin basal atau
aktivasi aksis HPA.125 Hiperaktivitas sistem saraf simpatik juga pasca OGTT. Meskipun metode ini dilakukan dalam kondisi
telah dilaporkan pada OSA.126 Pengobatan OSA dengan nasal fisiologis, tidak seperti teknik klem euglikemik, penulis melaporkan
continuous positive airway pressure secara istimewa menurunkan nilai r sebesar 0,93 hingga 0,99 untuk korelasi antar metode.132
jaringan adiposa viseral.127 Meskipun studi ini tidak
mengesampingkan resistensi insulin sebagai kontributor OSA, ada
beberapa mekanisme, yang dijelaskan di atas, dimana OSA Ukuran Fungsional Resistensi Insulin
mungkin berkontribusi terhadap resistensi insulin. Pendekatan lain adalah untuk mengidentifikasi pasien resisten
insulin, berdasarkan penanda fungsional resistensi insulin.
Pengukuran Insulin dan Resistensi Insulin McLoughlin et al mampu mengidentifikasi individu yang resisten
Ada berbagai pendekatan untuk penilaian laboratorium terhadap terhadap insulin dari kohort yang kelebihan berat badan dengan
resistensi insulin. Selama bertahun-tahun, kota spesifik yang melihat konsentrasi trigliserida plasma, rasio trigliserida terhadap
terbatas dari radio-immunoassay lama yang bereaksi silang dengan konsentrasi kolesterol lipoprotein densitas tinggi dan konsentrasi
proinsulin telah mengurangi kredibilitas pengukuran resistensi insulin. Dengan menggunakan titik potong 1,47 mmol/L untuk
insulin dalam pengaturan klinis. Tes saat ini telah meningkatkan trigliserida, 1,8 mmol/L untuk rasio kolesterol lipoprotein densitas
kota dan presisi tertentu. Tinjauan komprehensif tes insulin berada tinggi trigliserida dan 109 pmol/L (16 mIU/L) untuk insulin, mereka
di luar cakupan tinjauan ini dan pembaca didorong untuk mencapai sensitivitas dan spesifikasi yang sebanding dengan
berkonsultasi dengan Sapin128 dalam hal ini. Resistensi insulin Pengobatan Dewasa Panel III untuk mendiagnosa sindrom metabolik.133
dapat diukur dengan melihat langsung pengambilan glukosa yang
dimediasi insulin pada keadaan basal atau pascastimulasi, dengan Komentar Penutup Satu
menyimpulkan dari konsentrasi relatif glukosa dan insulin, atau abad atau lebih sejak penelitian di bidang ini dimulai dengan
dengan melihat penanda pengganti tindakan insulin. sungguh-sungguh, baik signifikansi maupun minat medis dan ilmiah
di bidang ini tidak berkurang. Alih-alih, globalisasi yang cepat,
Metode penelitian urbanisasi, dan industrialisasi telah melahirkan epidemi obesitas,
Metode yang paling diterima untuk mengukur resistensi insulin diabetes, dan penyakit penyerta yang menyertainya, karena
terbatas pada pengaturan penelitian karena invasi dan ketidakaktifan fisik dan makanan "kenyamanan" membuka tabir sifat
kompleksitasnya. Studi penjepit secara khusus mengukur genetik predisposisi laten. Mekanisme biologis rumit dan rumit dan
penyerapan glukosa yang dimediasi insulin dalam kondisi terkontrol. tidak sepenuhnya dipahami.
Ini termasuk klem euglikemik (dianggap sebagai standar emas) dan Namun, mundur selangkah, kita mungkin perlu mempertimbangkan
klem hiperinsulinemia. Kurang invasif adalah tes sensitivitas insulin perubahan sosial yang dramatis di abad yang lalu sehubungan
dan tes toleransi insulin pendek. Pelajari segera dengan aktivitas fisik, pola makan, pekerjaan, sosialisasi, dan pola tidur.
34 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
Selain tantangan yang tersisa dalam mengungkap faktor dari reseptor membran ke saluran nukleus. Diabetes.
genetik dan mekanistik, mungkin tantangan yang lebih besar 2004;53 Supl 1:S86-91.
adalah secara kreatif, untuk menyesuaikan gaya hidup 14. MacDonald PE, El-Kholy W, Riedel MJ, Salapatek AM, PE
kontemporer dengan susunan genetik dan kebutuhan fisiologis kita. Ringan, Wheeler MB. Berbagai tindakan GLP-1 pada
proses sekresi insulin yang distimulasi glukosa.
Kepentingan bersaing: tidak ada yang diumumkan. Diabetes. 2002;51 Supl 3:S434-42.
15. Havel PJ. Sinyal perifer yang menyampaikan informasi
Referensi metabolisme ke otak: pengaturan asupan makanan dan
homeostasis energi jangka pendek dan jangka panjang.
1.Organisasi Kesehatan Dunia. Obesitas: Mencegah dan Exp Bio Med (Maywood). 2001;226:963-77.
Mengelola Laporan Epidemi Global dari Seri Laporan 16. Nielsen JH, Serup P. Molekuler dasar pengembangan
Teknis Konsultasi WHO. Organisasi Kesehatan Dunia, pulau, pertumbuhan dan regenerasi. Opini Saat Ini dalam
Jenewa 2000. Endokrinologi & Diabetes 1998;5:97-107.
2. Cefalu WT. Resistensi insulin: konsep seluler dan klinis. Exp 17. Kahn SE, McCulloch DK, Porte D. Sekresi insulin pada
Bio Med (Maywood). 2001;226:13-26. manusia normal dan diabetes. Dalam: Alberti KGMM,
3. Reaven G. Sindrom metabolik atau sindrom resistensi Zimmet P, Defronzo RA, editor & Keen H, (hon) editor.
insulin? Beda nama, beda konsep, beda tujuan. Endocrinol Buku Teks Internasional Diabetes Mellitus. ( edisi ke-2)
Metab Clin North Am.2004;33:283-303. John Wiley & Sons, New York; 1997 hal. 337-54.
18. Chen M, Porte D Jr. Pengaruh kecepatan dan dosis infus
4. Bliss M. Sejarah Insulin Diabetes Care. 1993;16: Supl glukosa pada respon insulin akut pada manusia. J Clin
3:4-7. Endokrinol Metab. 1976;42:1168-75.
5.Rumah PD. Terapi insulin. Dalam: Alberti KGMM, Zimmet P, 19. Liu Z, Barrett EJ. Metabolisme protein manusia: pengukuran
editor Defronzo RA & editor Keen H (Hon) International dan pengaturannya. Am J Physiol Endocrinol Metab.
Textbook of Diabetes Mellitus ( edisi ke-2) 2002;283:E1105-12.
John Wiley & Sons, New York; 1997 hal. 899-928. 20. Amery CM, Nattrass M. Asam lemak dan sekresi insulin.
6. Dodson G, Steiner D. Peran perakitan dalam biosintesis Diabetes Obes Metab. 2000;2:213-21.
insulin. Curr Opin Struktur Biol. 1998;8:189-94. 21. Frost GS, Brynes AE, Dhillo WS, Bloom SR, McBurney MI.
7. Schroder D, Teknologi Zuhlke H. Gene, karakterisasi gen Efek pengayaan serat pasta dan kandungan lemak pada
insulin dan hubungannya dengan penelitian diabetes. pengosongan lambung, GLP-1, glukosa, dan respons
Endokrinologi. 1982;79:197-209. insulin terhadap makanan. Eur J Clin Nutr. 2003;57:293-8.
22.Reaven GM. Pengaruh perbedaan jumlah dan jenis diet
8. Malaise WJ. Biosintesis dan sekresi insulin in vitro.
Di dalam: Editor Alberti KGMM, Zimmet P, Defronzo RA karbohidrat pada glukosa plasma dan respon insulin pada
& Keen H (Hon). International Textbook of Diabetes manusia. Am J Clin Nutr. 1979;32:2568-78.
Mellitus ( edisi ke-2) John Wiley & Sons, New York; 1997 23. WoleverTM. Indeks glikemik. Diet Nutr Dunia Rev.
hal. 315-36. 1990;62:120-85.
9. Porksen N, Hollingdal M, Juhl C, Butler P, Veldhuis JD, 24. Efendic S, Portwood N. Tinjauan tentang hormon inkretin.
Schmitz O. Sekresi insulin berdenyut: deteksi, regulasi, Horm Metab Res 2004;36:742-6.
dan peran dalam diabetes. Diabetes. 2002;51 Supl 25. Kido Y, Nakae J, Accili D. Reseptor insulin dan target
1:S245-54. selulernya. J Clin Endokrinol Metab. 2001;86:972-9.
10. Bratanova-Tochkova TK, Cheng H, Daniel S, dkk. 26. Withers DJ, White M. Perspektif: Sistem pensinyalan insulin
Mekanisme pemicu dan augmentasi, kumpulan granula, - mata rantai umum dalam patogenesis diabetes tipe 2.
dan sekresi insulin bifasik. Diabetes 2002;51 Supl. Endokrinologi. 2000;141:1917-21.
1:S83-90.
27. Burks DJ, White MF. Protein IRS dan fungsi sel beta.
11. Nielsen JH, Galsgaard ED, Moldrup A, dkk.Pengaturan Diabetes. 2001;50 Supl 1:S140-5.
massa sel beta oleh hormon dan faktor pertumbuhan
28. Hunter SJ, Garvey WT. Aksi insulin dan resistensi insulin:
Diabetes. 2001;50 Supl 1:S25-9.
penyakit yang melibatkan kerusakan pada reseptor
12. De Lonlay, Saudubray JM. Hipoglikemia hiperinsulinemia insulin, transduksi sinyal, dan sistem efektor transpor
persisten. Dalam: Fernandes J, Sudubray JM, editor van glukosa. Am J Med. 1998;105:331-45.
den Berghe Penyakit Metabolik Bawaan: Diagnosis dan 29. Karam JH. Hormon Pankreas dan Diabetes Mellitus.
pengobatan. ( edisi ke-3): Springer, Heidelberg Jerman;
Di dalam: Greenspan FS, Strewler GJ, editor. Endokrinologi
2000 hlm.117-26.
Dasar dan Klinis. Appleton & Lange, Stamford CT USA;
13. Soria B, Quesada I, Ropero AB, Pertusa JA, Martin F, 1997 hal. 601-2.
Nadal A. Pemain baru di pensinyalan pulau pankreas:
30. Denton RM, Tavare JM. Basis molekuler atau insulin
Klinik Biochem Rev Vol 26 Mei 2005 I 35
Machine Translated by Google
WilcoxG
tindakan pada metabolisme intraseluler. Di dalam: Alberti KGMM, implikasi. Histol Histopatol 1998;13:1215-24.
Zimmet P, Defronzo RA, Keen H (Hon), editor. 45. Unger JW, reseptor Lange W. Insulin di kelenjar hipofisis: bukti
International Textbook of Diabetes Mellitus ( edisi ke-2) morfologis pengaruh pada sel sintesis peptida opioid. Jaringan
John Wiley & Sons, New York; 1997 hal. 469-88. Sel Res 1997;288:471-83.
31. Clemmons DR. Analisis struktural dan fungsional faktor pertumbuhan
seperti insulin. Sdr. Med Bull. 1989;45:465-80. 46. Nagasaka Y, Kaneko T. [Biologi molekuler pengaturan struktur
fungsi ginjal, fungsi dan distribusi reseptor-insulin, glukagon]
32. Frystyk J, Ørskof H. IGFI, IGFII, protein pengikat IGF dan diabetes. Nippon Rinsho 1992;50:2921-4.
Di dalam: Alberti KGMM, Zimmet P, Defronzo RA, Keen H (hon),
editor. International Textbook of Diabetes Mellitus ( edisi ke-2) 47. Sechi LA, Bartoli E. Mekanisme molekuler resistensi insulin pada
John Wiley & Sons, New York; 1997 hal. 417-36. hipertensi arteri. Darah Pres Suppl 1996;1:47-54.
33. Wheatcroft SB, Williams IL, Shah AM, Kearney MT. 48. Poretsky L, Kalin MF. Fungsi gonadotropin insulin Endocre Rev
Implikasi patofisiologi resistensi insulin pada fungsi endotel 1987;8:132-41.
vaskular. Obat Diabetes. 2003;20:255-68.
49. Samoto T, Maruo T, Katayama K, Barnea ER, Mochizuk M.
Perubahan ekspresi insulin dan reseptor faktor pertumbuhan
34. Smith U. Gangguan pensinyalan dan aksi insulin ('diabetes') terjadi seperti insulin-1 di kompartemen folikel dan stroma ovarium
pada sel-sel lemak jauh sebelum intoleransi glukosa - apakah polikistik. Endocr J 1993;40:413-24.
resistensi insulin dimulai pada jaringan adiposa? Int J Obes Relat
50. Abele V, Pelletier G, Tremblay RR. Lokalisasi radioautografi dan
Metab Disord. 2002;26:897-904.
pengaturan reseptor insulin pada testis tikus. Resep J Res
35. Giorgino F, Laviola L, Eriksson JW. Perbedaan regional aksi insulin 1986;6:461-73.
dalam jaringan adiposa: wawasan dari studi in vivo dan in vitro.
51.Thomas DM, Udagawa N, Hards DK, dkk Ekspresi reseptor insulin
Pemindaian Acta Physiol. 2005;183:13-
dalam sel seperti oseoklas primer dan biakan. Tulang
30.
1998;23:181-6.
36. Halvatsiotis PG, Turk D, Alzaid A, Dinneen S, Rizza RA, Nair KS.
52. Lichtenstein AH, Schwab AS. Hubungan lemak makanan dengan
Efek insulin pada kinetika leusin pada diabetes melitus tipe 2.
metabolisme glukosa. Aterosklerosis. 2000;150:227-43.
Diabetes Nutr Metab. 2002;15:136-42.
53. Bray GA, Lovejoy JC, Smith SR, dkk Pengaruh Lemak dan Asam
37.Grundy SM. Apa kontribusi obesitas terhadap sindrom metabolik?
Lemak Berbeda pada Obesitas, Resistensi Insulin dan
Endocrinol Metab Clin North Am.2004;33:267-82.
Peradangan J. Nutr. 2002;132:2488-91.
38. Krauss RM, Siri PW. Kelainan metabolik: trigliserida dan lipoprotein
54. Sampath H, Ntambi JM. Regulasi asam lemak tak jenuh ganda dari
densitas rendah. Endocrinol Metab Clin North Am.2004;33:405-15.
ekspresi gen. Kacang Pdt. 2004;62:333-9.
39. Devaraj S, Rosenson RS, Jialal I. Sindrom metabolik: penilaian
55. Simopoulos AP. Asam lemak esensial dalam kesehatan dan
status pro-inflamasi dan prokoagulan. Endocrinol Metab Clin
penyakit kronis. Am J Clin Nutr. 1999 ;70(3 Suppl):560S-569S.
North Am.2004;33:431-53.
56. Borkman M, Chisholm DJ, Furler SM, dkk.Pengaruh suplementasi
minyak ikan terhadap metabolisme glukosa dan lipid pada
40. Stockhorst U, de Fries D, Steingrueber HJ, Scherbaum WA. Insulin
NIDDM. Diabetes. 1989;38:1314-9.
dan SSP: efek pada asupan makanan, memori, dan parameter
endokrin dan peran pemberian insulin intranasal pada manusia. 57. Friedberg CE, Janssen MJ, Heine RJ, Grobbee DE. Minyak ikan
Physiol Behav 2004;83:47-54. dan kontrol glikemik pada diabetes. Analisis meta.
Perawatan diabetes. 1998;21:494-500.
41. Gerozisis K. Insulin otak dan pemberian makan: komunikasi dua 58. WoleverTM. Karbohidrat makanan dan aksi insulin pada manusia.
arah. Eur J Pharmacol 2004:490:59-70. Br J Nutr. 2000;83 Supl 1:S97-102.
42. Watson GS, Craft S. Peran resistensi insulin dalam patogenesis 59. Toida S, Takahashi M, Shimizu H, Sato N, Shimomura Y, Kobayashi
penyakit Alzheimer: implikasi untuk pengobatan. Obat CNS I. Pengaruh pemberian sukrosa tinggi pada akumulasi lemak
pada tikus Wistar jantan. Obes Res 1996;4:561-8.
2003;17:27-45.
43.Kulkarni RN. Sel pembayaran pulau kecil. Sel Int J Biochem
Biol. 2004;36:365-71. 60. Davy BM, Melby CL. Efek karbohidrat kaya serat pada fitur
Syndrome X. J Am Diet Assoc. 2003;103:86-96.
44. Unger JW, reseptor Betz M. Insulin dan protein transduksi sinyal
dalam sistem hipotalamo-hipofisis: ulasan tentang temuan
morfologis dan fungsional 61. Beck B, Villaume C, Bau HM, dkk.Pengaruh jangka panjang dari
diet suplemen dedak gandum pada sekresi
36 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
hormon gastrointestinal dan penyerapan nutrisi pada pria sehat. 77. Tuomilehto J, Lindstrom J, Eriksson JG, et al.Kelompok Studi
Hum Nutr Clin Nutr. 1986;40:25-33. Pencegahan Diabetes Finlandia.. Pencegahan diabetes melitus
62. Cani PD, Dewever C, Delzenne NM. Fruktan tipe inulin memodulasi tipe 2 dengan perubahan gaya hidup di antara subjek dengan
peptida gastrointestinal yang terlibat dalam pengaturan nafsu gangguan toleransi glukosa. N Engl J Med 2001;344:1343-50.
makan (peptida-1 seperti glukagon dan ghrelin) pada tikus.
Br J Nutr. 2004;92:521-6. 78.Dohm GL. Regulasi ekspresi GLUT-4 otot rangka dengan olahraga.
63. Roberfroid M. Serat makanan, inulin, dan oligofruktosa: ulasan yang J Appl Physiol 2002:93:782-7.
membandingkan efek fisiologisnya. Crit Rev Food Sci Nutr. 79. Henriksen EJ. Efek latihan akut dan latihan olahraga pada resistensi
1993;33:103-48. . Erratum di: Crit Rev Food Sci Nutr 1993;33:553. insulin. J Appl Physiol. 2002; 93:788-96.
64. Higgins YA. Pati resisten: efek metabolisme dan potensi manfaat 80. Juleen R.Zierath. Latihan olahraga menginduksi perubahan dalam
kesehatan. J AOAC Int. 2004;87:761-8. pensinyalan insulin pada otot rangka. J Appl Physiol
65. Linn T, Santosa B, Gronemeyer D, dkk Pengaruh asupan protein 2002;93:773-81.
diet jangka panjang pada metabolisme glukosa pada manusia. 81. Van den Berghe G. Bagaimana kontrol glukosa darah dengan insulin
Diabetologi. 2000;43:1257-65. menyelamatkan nyawa dalam perawatan intensif? Investasi J
66. Greenfield JR, Samaras K, Hayward CS, Chisholm DJ, Campbell Clin. 2004;114:1187-95.
LV. Efek postprandial yang menguntungkan dari sejumlah kecil 82. Marette A. Mediator resistensi insulin yang diinduksi sitokin pada
alkohol pada diabetes dan faktor risiko kardiovaskular: Modifikasi obesitas dan pengaturan inflamasi lainnya.
oleh Resistensi Insulin. J Clin Curr Opin Clin Nutr Metab Care 2002;5:377-83.
Endokrinol Metab. 2 Nov 2004 [Epub sebelum dicetak] 83. Rosmond R. Peran stres dalam patogenesis sindrom metabolik.
67. Magis DC, Jandrain BJ, Scheen AJ. Alkohol, sensitivitas insulin dan Psikoneuroendokrinologi. 2005;30:1-10.
diabetes. Pdt Med Liege. 2003;58:501-7.
68.Anderson RA. Kromium, intoleransi glukosa dan diabetes.J Am Coll 84. Spiegel K, Leproult R, Van Cauter E. Dampak hutang tidur pada
Nutr. 1998;17:548-55. fungsi metabolisme dan endokrin. Lanset. 1999;354:1435-9.
69.Vincent JB. Biokimia kromium. J Nutr. 2000;130:715-8.
85. Vgontzas AN, Mastorakos G, Bixler EO, Kales A, Gold PW,
70. Fernandez-Real JM, Lopez-Bermejo A, Ricart W. Chrousos GP. Efek kurang tidur pada aktivitas sumbu hipotalamus-
Pembicaraan silang antara metabolisme zat besi dan diabetes. hipofisis-adrenal dan pertumbuhan: implikasi klinis potensial.
Diabetes. 2002;51:2348-54. Klinik Endokrinol (Oxf). 1999;51:205-15.
71. Bhathena SJ, Velasquez MT. Peran menguntungkan fitoestrogen
diet pada obesitas dan diabetes. 86. Vorona RD, Winn MP, Babineau TW, Eng BP, Feldman HR, Ware
Am J Clin Nutr. 2002;76:1191-201. JC. Pasien yang kelebihan berat badan dan obesitas pada
populasi perawatan primer melaporkan kurang tidur dibandingkan
72. Broadhurst CL, Polansky MM, Anderson RA. Insulin seperti aktivitas
pasien dengan indeks massa tubuh normal.
biologis ekstrak air tanaman kuliner dan obat in vitro. Kimia
Arch Intern Med. 2005; 165:25-30.
Makanan J Agric. 2000;48:849-52.
87. Spiegel K, Tasali E, Penev P, Van Cauter E. Pembatasan tidur pada
pria muda yang sehat dikaitkan dengan penurunan kadar leptin,
73. Grassi D, Lippi C, Necozione, Desideri, Ferri C. Pemberian dark
peningkatan kadar ghrelin, dan peningkatan rasa lapar dan nafsu
chocolate jangka pendek diikuti oleh peningkatan sensitivitas
makan. Ann Intern Med. 2004;141:846-50.
insulin yang signifikan dan penurunan tekanan darah pada orang
sehat. Am J Clin Nutr 2005;81:611-4.
88. Mullington J, Hermann D, Holsboer F, Pollmacher T.
Penekanan kadar hormon pertumbuhan nokturnal yang
74. Tomas E, Zorzano A, Ruderman NB. Latihan dan pensinyalan
bergantung pada usia selama kurang tidur. Neuroendokrinologi.
insulin: perspektif sejarah. J Appl Physiol. 2002;93:765-72.
1996;64:233-41.
89. Hew FL, O'Neal D, Kamarudin N, Alford FP, JD Terbaik.
75. Manson JE, Nathan DM, Krolewski AS, Stampfer MJ, Willett WC,
Defisiensi hormon pertumbuhan dan risiko kardiovaskular.
Hennekens CH. Sebuah studi prospektif tentang olahraga dan
Baillieres Clin Endocrinol Metab. 1998; 12:199-216.
kejadian diabetes di antara dokter pria AS. JAMA. 1992;268:63-7.
90. Tassone F, Lanfranco F, Gianotti L, dkk Sindrom apnea tidur
obstruktif merusak sensitivitas insulin secara independen dari
76. Knowler WC, Barrett-Connor E, Fowler SE, dkk.Kelompok Riset
variabel antropometrik. Klinik Endokrinol (Oxf). 2003;59:374-9.
Program Pencegahan Diabetes.
Pengurangan kejadian diabetes tipe 2 dengan intervensi gaya
hidup atau metformin. NEJM 2002;346:393-403. 91. Yee B, Liu P, Philips C, Grunstein R. Perubahan neuroendokrin
pada sleep apnea. Curr Opin Pulm Med.
Klinik Biochem Rev Vol 26 Mei 2005 I 37
Machine Translated by Google
WilcoxG
2004;10:475-81. diabetes: mekanisme molekuler dan presentasi klinis. Diabetes.
92. Butte NF. Metabolisme karbohidrat dan lipid pada kehamilan: 2004;53 Supl 1:S103-9.
normal dibandingkan dengan diabetes melitus gestasional. Am 108. Panetta J, Smith LJ, Boneh A. Pengaruh vitamin dosis tinggi,
J Clin Nutr 2000;71(suppl):1256S-61S. koenzim Q dan diet tinggi lemak pada pasien anak dengan
93. Seely EW, dan Solomon CG. Resistensi insulin dan peran penyakit mitokondria. J Mewarisi Metab Dis. 2004;27:487-98.
potensinya dalam hipertensi yang diinduksi kehamilan. J Clin
Endokrinol Metab. 2003;88:2393-8. 109. Pedersen KF, Dufour S, Befroy D, Garcia R, Shulman GI.
94. Samar-samar J. Tingkat diferensiasi maskulin obesitas: faktor yang Gangguan aktivitas mitokondria pada keturunan resisten insulin
menentukan predisposisi diabetes, aterosklerosis, asam urat, dari pasien dengan diabetes tipe 2. N Engl J Med 2004;350:660-71.
dan penyakit batu urat. 1956.
Obes Res. 1996;4:204-12. 110. Giacchero R, Fiorir L, Scaglioni S, Sala M, Giovanni M. Resistensi
95. Aronne LJ, Segal KR. Ukuran hasil distribusi adipositas dan lemak: insulin dan fungsi pankreas pada penyakit penyimpanan glikogen
penilaian dan implikasi klinis. Tipe 1. J Mewarisi Metab Dis 2003;26(suppl 2):124.
Obes Res. 2002;10 Tambahan 1:14S-21S.
96. Tomlinson JW, Sinha B, Bujalska I, Hewison M, Stewart PM. 111. Scheuner MT, Raffel LJ, Rotter JI. Genetika Diabetes.
Ekspresi 11beta-hidroksisteroid dehidrogenase tipe 1 di jaringan Di dalam: Alberti KGMM, Zimmet P, Defronzo RA, Keen H (hon),
adiposa tidak meningkat pada obesitas manusia. editor. International Textbook of Diabetes Mellitus ( edisi ke-2)
J Clin Endokrinol Metab. 2002;87:5630-5. John Wiley & Sons, New York; 1997 hal. 37-88.
97. Perseghin G, Petersen K, Shulman GI. Mekanisme seluler resistensi
insulin: hubungan potensial dengan peradangan. Int J Obes 112. Bornstein J. Sebuah teknik untuk pengujian sejumlah kecil insulin
Relat Metab Disord. 2003;27 Dlm 3:S6-11. menggunakan aloksan tikus diabetes, hypophysectomized,
adrenalectomized. Aust J Exp Biol Med Sci. 1950; 28:87-91.
98. Jansson PA, Pellme F, Hammarstedt A, et al.Sebuah penanda
seluler novel resistensi insulin dan aterosklerosis dini pada 113. Yalow, RS, Berson, SA Immunoassay insulin plasma endogen
manusia terkait dengan gangguan diferensiasi sel lemak dan pada manusia. J.Clin. menginvestasikan. 1960;39:1157-75.
adiponektin rendah. FASEB J. 2003;17:1434-40.
114. Bjorntorp P. Apakah reaksi stres menyebabkan obesitas perut
99.Reaven GM. Pentingnya mengidentifikasi pasien kelebihan berat dan penyakit penyerta? Obes Pdt. 2001;2:73-86.
badan yang paling diuntungkan dengan menurunkan berat 115. Bass J, Turek FW. Kurang Tidur di Amerika: Jalan Menuju
badan. Ann Intern Med. 2003;138:420-3. Obesitas dan Sindrom Metabolik Arch Intern Med. 2005;165:15-6.
100. Bressler P, De Fronzo RA. Di dalam: Alberti KGMM, Zimmet P,
Defronzo RA, Keen H (hon), editor. International Textbook of 116. Kotor SM. Apa kontribusi obesitas terhadap sindrom metabolik?
Diabetes Mellitus ( edisi ke-2) John Wiley & Sons, New York; Endocrinol Metab Clin North Am.2004;33:267-82.
1997 hal. 213-54.
101. Ananth J, Parameswaran S, Gunatilake S. Efek samping obat 117.Reaven GM. Resistensi insulin/hiperinsulinemia kompensasi,
antipsikotik atipikal. Curr Pharma Des. 2004;10:2219-29. hipertensi esensial, dan penyakit kardiovaskular. J Clin
Endokrinol Metab. 2003;88:2399-403.
102. Chen D, Misra A, Garg A. Lipodystrophy pada pasien yang
terinfeksi virus human immunodeficiency. J Clin Endokrinol 118. Wang CC, Goalstone ML, Draznin B. Mekanisme molekuler
Metab. 2002;87:4845-56. resistensi insulin yang memengaruhi biologi kardiovaskular.
103. Lebovitz HE. Agen antidiabetes oral. Med Clinic North Diabetes. 2004;53:2735-40.
Am.2004;88:847-63, ix-x. 119. Shinozaki K, Kashiwagi A, Masada M, Okamura T.
104. Ehrmann DA. Sindrom ovarium polikistik. N Engl J Med. Mekanisme molekuler gangguan fungsi endotel terkait dengan
2005;352:1223-36. resistensi insulin. Obat Curr Menargetkan Gangguan Hematol
Kardiovaskular. 2004;4:1-11.
105. Fernandez J, IPK Smith. Penyakit penyimpanan glikogen.
Di dalam: Fernandes J, Sudubray JM, van den Berghe, editor. 120. Jacobs HS, Conway GS. Leptin, ovarium polikistik dan sindrom
Penyakit Metabolik Bawaan: Diagnosis dan Perawatan. ( edisi ovarium polikistik. Pembaruan Hum Reprod. 1999;5:166-71.
ke-3). Springer, Heidelberg Jerman; 2000 hal. 86-101.
121. Angulo P, Lindor KD. Penyakit hati berlemak nonalkohol.
106. Garg A, Misra A Lipodystrophies: kelainan langka yang J Gastroenterol Hepatol. 2002;17 Suppl:S186-90.
menyebabkan sindrom metabolik. Endocrinol Metab Clin North 122. Calle EE, Kaaks R. Kelebihan berat badan, obesitas dan kanker:
Am.2004;33:305-31. bukti epidemiologis dan mekanisme yang diusulkan.
107. Maassen JA, Hart LM, Van Essen E, dkk.Mitokondria Kanker Rev Basah. 2004;4:579-91.
38 I Klinik Biochem Rev Vol 26 Mei 2005
Machine Translated by Google
Insulin dan Resistensi Insulin
123. Komninou D, Ayonote A, Richie JP Jr, Rigas B. Resistensi insulin
dan kontribusinya terhadap karsinogenesis usus besar.
Exp Bio Med (Maywood). 2003;228:396-405.
124. Kaaks R. Nutrisi, hormon, dan kanker payudara: apakah insulin
merupakan mata rantai yang hilang? Pengendalian Penyebab
Kanker. 1996;7:605-25.
125. Lanfranco F, Gianotti L, Pivetti S, et al.Pasien obesitas dengan
sindrom apnea tidur obstruktif menunjukkan perubahan yang khas
pada kortikotrof tetapi tidak pada fungsi tirotrof dan laktotrof. Klinik
Endokrinol (Oxf). 2004;60:41-8.
126. Wilcox I, McNamara SG, Collins FL, Grunstein RR, Sullivan CE.
"Syndrome Z": interaksi sleep apnea, faktor risiko vaskular dan
penyakit jantung. Dada.
1998;53 Dlm 3:S25-8.
127. Chin K, Shimizu K, Nakamura T, et al.Perubahan lemak visceral
intraabdomen dan kadar leptin serum pada pasien dengan
sindrom apnea tidur obstruktif setelah terapi tekanan saluran
napas positif kontinyu hidung. sirkulasi. 1999;100:706-12.
128. Uji Sapin R. Insulin: fitur analitik yang diketahui sebelumnya dan
baru. Laboratorium Klinik. 2003;49:113-21.
129. Wallace TM, Retribusi JC, Matthews DR. Penggunaan dan
penyalahgunaan pemodelan HOMA. Perawatan Diabetes 2004;27:1487.
130. Wallace TM, Matthews DR. Penilaian resistensi insulin pada pria.
Diabetes Med 2002;19:527-34.
131. Sikaris KA. Biokimia klinis obesitas. Klinik
Biochem Rev 2004;25:165-82.
132. Belfiore F, Ianello S, Volpicelli G. Indeks sensitivitas insulin dihitung
dari kadar insulin, glukosa, dan FFA yang diinduksi OGTT dan
basal. Mol Genet Metab 1998;63:134-41.
133. McLaughlin T, Abbasi F, Cheal K, Chu J, Lamendola C, Reaven G.
Penggunaan penanda metabolik untuk mengidentifikasi individu
kelebihan berat badan yang resisten terhadap insulin. Ann Intern
Med. 2003;139:802-9.
Klinik Biochem Rev Vol 26 Mei 2005 I 39
Lihat statistik publikasi
Anda mungkin juga menyukai
- Referat DM Pada AnakDokumen47 halamanReferat DM Pada AnakKhansadhia Hasmaradana Mooiindie DjojonegoroBelum ada peringkat
- Mechanisms of Insulin Action and Insulin Resistance - PMCDokumen201 halamanMechanisms of Insulin Action and Insulin Resistance - PMCyz2bg7xf4zBelum ada peringkat
- Tinjauan Molekular Dan Aspek Klinis Resistensi InsulinDokumen8 halamanTinjauan Molekular Dan Aspek Klinis Resistensi InsulinAsiah JelitaBelum ada peringkat
- Resistensi Insulin (Referat)Dokumen14 halamanResistensi Insulin (Referat)Ai AiiBelum ada peringkat
- RESISTENSI INSULIN Jurnal ReadingDokumen14 halamanRESISTENSI INSULIN Jurnal ReadingAnfasah KimikoBelum ada peringkat
- G12der - BAB II Tinjauan PustakaDokumen6 halamanG12der - BAB II Tinjauan PustakaMPMSarah Nadya WiliyaniBelum ada peringkat
- Resistensi InsulinDokumen34 halamanResistensi InsulinAnonymous fOcbAtBelum ada peringkat
- Peran Insulin Dalam Kesehatan Dan Penyakit - Pembaruan - PMCDokumen32 halamanPeran Insulin Dalam Kesehatan Dan Penyakit - Pembaruan - PMCyz2bg7xf4zBelum ada peringkat
- Resistensi Insulin Pada KehamilanDokumen26 halamanResistensi Insulin Pada KehamilanZulfahmi ZulfaBelum ada peringkat
- Elsina Irianti Pahabol - LKM Skenario 3 Blok EndokrinDokumen23 halamanElsina Irianti Pahabol - LKM Skenario 3 Blok Endokrinelzhy 12Belum ada peringkat
- Patofisiologi Diabetes MellitusDokumen12 halamanPatofisiologi Diabetes MellitusBram Wongso100% (1)
- AdiponektinDokumen7 halamanAdiponektinAnastasia Okta ErishaBelum ada peringkat
- REKAYASA GENETIKA INSULIN DR - HegiDokumen22 halamanREKAYASA GENETIKA INSULIN DR - HegiHegi DarmawanBelum ada peringkat
- Askep Diabetes MellitusDokumen19 halamanAskep Diabetes Mellitus3 raBelum ada peringkat
- Template Literature Review Sa12Dokumen3 halamanTemplate Literature Review Sa12astrimillianiBelum ada peringkat
- LP DM YulianiDokumen29 halamanLP DM YulianiNi Kadek Dwi ApriantiBelum ada peringkat
- PBL Skenario 1 B7Dokumen38 halamanPBL Skenario 1 B7farhanBelum ada peringkat
- Dom0013-0677 en IdDokumen8 halamanDom0013-0677 en IdBelia Bima NafisaBelum ada peringkat
- Wrap Up Skenario 1 EndokrinDokumen30 halamanWrap Up Skenario 1 EndokrinJoko WijanarkoBelum ada peringkat
- Hormon Insulin Dan GlukagonDokumen49 halamanHormon Insulin Dan GlukagonAan AndriyansirBelum ada peringkat
- LP Eka Saputri Minggu 1Dokumen34 halamanLP Eka Saputri Minggu 1wanaa0806Belum ada peringkat
- Tugas KMB Eka Saputri Minggu Ke 1Dokumen69 halamanTugas KMB Eka Saputri Minggu Ke 1wanaa0806Belum ada peringkat
- Laporan Pendahuluan - DM Novi NursifaDokumen24 halamanLaporan Pendahuluan - DM Novi NursifaNovi NursifaBelum ada peringkat
- Konsep DMDokumen35 halamanKonsep DMAnang SetyaBelum ada peringkat
- Bioteknologi Modern Pembuatan Hormon Insulin FIXXDokumen16 halamanBioteknologi Modern Pembuatan Hormon Insulin FIXXKillua HanterBelum ada peringkat
- SK 1 B-13 YarsiDokumen36 halamanSK 1 B-13 YarsiIyad PabottingiBelum ada peringkat
- InsulinDokumen7 halamanInsulinFeni ArmadaniBelum ada peringkat
- Asuhan Keperawatan Pada Pasien Dengan Diabetes MellitusDokumen97 halamanAsuhan Keperawatan Pada Pasien Dengan Diabetes MellitusPusat Layanan Kesehatan IAIN PontianakBelum ada peringkat
- Bioteknologi Insulin-1Dokumen7 halamanBioteknologi Insulin-1Misa RaemaBelum ada peringkat
- Peran Adiponektin Terhadap Kejadian Resistensi Insulin Pada Sindrom Metabolik 4Dokumen6 halamanPeran Adiponektin Terhadap Kejadian Resistensi Insulin Pada Sindrom Metabolik 4Husni IrchyBelum ada peringkat
- Askep DM Pada AnakDokumen30 halamanAskep DM Pada Anakaliyah mahmudah100% (2)
- Laporan Pendahuluan Taupik Muslim DMDokumen28 halamanLaporan Pendahuluan Taupik Muslim DMDinda AyundaBelum ada peringkat
- Referat 3 Tari PPDSDokumen17 halamanReferat 3 Tari PPDSRahma Evasari LubisBelum ada peringkat
- DM Nurul MDokumen29 halamanDM Nurul MNurul Mutmainnah SyukurBelum ada peringkat
- Materi InsulinDokumen4 halamanMateri InsulinHastrilia BuntangBelum ada peringkat
- Tugas Kelompok 4 Konsep Asuhan Keperawatan Komunitas Pada Agregat Kelompok Penyakit Diabetes MelitusDokumen23 halamanTugas Kelompok 4 Konsep Asuhan Keperawatan Komunitas Pada Agregat Kelompok Penyakit Diabetes MelitusDINDABelum ada peringkat
- Boisel Muhammad RidhanaDokumen6 halamanBoisel Muhammad RidhanaMuhammad RidhaBelum ada peringkat
- Referat Leaky Gut Syndrome AndrewDokumen15 halamanReferat Leaky Gut Syndrome AndrewAndrew HartonoBelum ada peringkat
- Anatomi Fisiologi PankreasDokumen25 halamanAnatomi Fisiologi PankreasYesti Toree100% (1)
- Makalah KMB II Kel 2Dokumen36 halamanMakalah KMB II Kel 2YulitaBelum ada peringkat
- Laporan Pendahuluan DMDokumen28 halamanLaporan Pendahuluan DMFenti GuspitaBelum ada peringkat
- LP Ulkus DiabetikumDokumen18 halamanLP Ulkus DiabetikumAnnisaBelum ada peringkat
- LP Sifera Elva (Diabetes Melitus)Dokumen34 halamanLP Sifera Elva (Diabetes Melitus)Rima WulandariBelum ada peringkat
- Stase Keperawatan Medikal Bedah (KMB)Dokumen22 halamanStase Keperawatan Medikal Bedah (KMB)florentina lulutBelum ada peringkat
- Pemicu 3 Endokrin - Kel. 19 - 20Dokumen45 halamanPemicu 3 Endokrin - Kel. 19 - 20zahfira edwardBelum ada peringkat
- Juvenile DiabetesDokumen12 halamanJuvenile DiabetesIndri DwiBelum ada peringkat
- BAB I, 2,3 Referat Terapi InsulinDokumen17 halamanBAB I, 2,3 Referat Terapi Insulindbkyh100% (1)
- RekombinantDokumen5 halamanRekombinanttri sutriani syamBelum ada peringkat
- Hormon InsulinDokumen18 halamanHormon InsulinNur AzizahBelum ada peringkat
- LP DMDokumen15 halamanLP DMHerman PitayadiBelum ada peringkat
- LP Diabetes MelitusDokumen22 halamanLP Diabetes MelitusNalau Sapu RataBelum ada peringkat
- Fisiologi Kerja InsulinDokumen4 halamanFisiologi Kerja InsulinPatricia KalangiBelum ada peringkat
- Makalah Insulin: Bab I PendahuluanDokumen14 halamanMakalah Insulin: Bab I PendahuluanMeliyana hayatiBelum ada peringkat
- Made Adhitya (LP DM)Dokumen34 halamanMade Adhitya (LP DM)Novi MahritaBelum ada peringkat
- HestiDokumen58 halamanHestiBungaBelum ada peringkat
- Panduan Esensial untuk Skoliosis dan Kesehatan KehamilanDari EverandPanduan Esensial untuk Skoliosis dan Kesehatan KehamilanPenilaian: 4.5 dari 5 bintang4.5/5 (4)
- Jurnal Perawatan Skoliosis Natural Anda: Petunjuk per hari selama 12 minggu untuk tulang belakang yang lebih lurus dan kuat!Dari EverandJurnal Perawatan Skoliosis Natural Anda: Petunjuk per hari selama 12 minggu untuk tulang belakang yang lebih lurus dan kuat!Penilaian: 4.5 dari 5 bintang4.5/5 (7)
- ARTHROLOGIDokumen14 halamanARTHROLOGIKhansa RosviBelum ada peringkat
- INSULINNDokumen7 halamanINSULINNKhansa RosviBelum ada peringkat
- Sistem Saraf PusatDokumen14 halamanSistem Saraf PusatKhansa RosviBelum ada peringkat
- Sitoplasma: SitosolDokumen11 halamanSitoplasma: SitosolKhansa RosviBelum ada peringkat
- Siklus SelDokumen6 halamanSiklus SelKhansa RosviBelum ada peringkat
- Peran Dan Fungsi Hospital by LawsDokumen2 halamanPeran Dan Fungsi Hospital by LawsKhansa RosviBelum ada peringkat
- Anatomi 1Dokumen3 halamanAnatomi 1Khansa RosviBelum ada peringkat
- Doctor Patient RelationshipDokumen3 halamanDoctor Patient RelationshipKhansa RosviBelum ada peringkat
- Dasar Dasar ProfesionalismeDokumen4 halamanDasar Dasar ProfesionalismeKhansa RosviBelum ada peringkat
- Praktik KedokteranDokumen4 halamanPraktik KedokteranKhansa RosviBelum ada peringkat
- Mac ConkeyDokumen4 halamanMac ConkeyKhansa RosviBelum ada peringkat
- Tugas Bahasa IndonesiaDokumen4 halamanTugas Bahasa IndonesiaKhansa RosviBelum ada peringkat
- Skala LikertDokumen4 halamanSkala LikertKhansa RosviBelum ada peringkat
- Skala Likert Adalah Skala Yang Digunakan Untuk Mengukur PersepsiDokumen3 halamanSkala Likert Adalah Skala Yang Digunakan Untuk Mengukur PersepsiKhansa RosviBelum ada peringkat
- Definisi Pola MakanDokumen3 halamanDefinisi Pola MakanKhansa Rosvi100% (1)
- Pola MakanDokumen2 halamanPola MakanKhansa RosviBelum ada peringkat