Anda mungkin juga menyukai
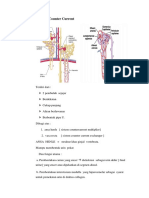
- Mekanisme Counter CurrentDokumen5 halamanMekanisme Counter CurrentLeticiaBelum ada peringkat
- ASKEP DIABETESDokumen21 halamanASKEP DIABETESNufie Crunch Chic75% (4)
- Hormon Yang Berpengaruh Di System UrinariaDokumen6 halamanHormon Yang Berpengaruh Di System UrinariaQurotulAqyunBelum ada peringkat
- Proses Terjadinya Pengenceran Dan Pemekatan UrineDokumen3 halamanProses Terjadinya Pengenceran Dan Pemekatan UrineCho Maharani Rijhigo Bae100% (2)
- OSMOLARITASDokumen8 halamanOSMOLARITASAneu Nurdiana100% (4)
- Fisiologi HausDokumen4 halamanFisiologi HausditanhBelum ada peringkat
- KafeinDokumen11 halamanKafeinputridwierlinda100% (1)
- FISIOREGULASI URINDokumen2 halamanFISIOREGULASI URINNafiys Reeven TeergovBelum ada peringkat
- Pengaturan Produksi ADHDokumen23 halamanPengaturan Produksi ADHWindryTandibunnaBelum ada peringkat
- Ringkasan Mekanisme Pemekatan Urin Dan Perubahan Osmolaritas Di Berbagai Segmen TubulusDokumen3 halamanRingkasan Mekanisme Pemekatan Urin Dan Perubahan Osmolaritas Di Berbagai Segmen TubulusBalqis DwiyantiBelum ada peringkat
- REGULASI REABSORPSI H20 OLEH VASOPRESINDokumen4 halamanREGULASI REABSORPSI H20 OLEH VASOPRESINAnnisaBelum ada peringkat
- PengantarDokumen5 halamanPengantarauliya nidaBelum ada peringkat
- PERAN GINJAL DALAM HOMEOSTASISDokumen4 halamanPERAN GINJAL DALAM HOMEOSTASISfaradila_cahyaniBelum ada peringkat
- AMI-Pengaturan Osmolaritas CES Dan Konsentrasi NatriumDokumen43 halamanAMI-Pengaturan Osmolaritas CES Dan Konsentrasi NatriumMary AmellBelum ada peringkat
- Hormon Regulasi Cairan TubuhDokumen6 halamanHormon Regulasi Cairan TubuhMutia AgustriaBelum ada peringkat
- B5 - Ex 9 Act 6Dokumen17 halamanB5 - Ex 9 Act 6salma najmiBelum ada peringkat
- Soal-Soal Tentang Osmoregulasi Dan EkskresiDokumen8 halamanSoal-Soal Tentang Osmoregulasi Dan EkskresiSyarif Hidayat AmrullahBelum ada peringkat
- HORMON ADH MENYELESAIKAN TUGASNYADokumen12 halamanHORMON ADH MENYELESAIKAN TUGASNYAUlfah Rufaidah100% (1)
- OSMOSIS DAN REGULASI URINDokumen15 halamanOSMOSIS DAN REGULASI URINrara prameiBelum ada peringkat
- OPTIMASI FUNGSI GINJALDokumen30 halamanOPTIMASI FUNGSI GINJALRefanAldoBelum ada peringkat
- Fisiologi GinjalDokumen8 halamanFisiologi GinjalAmanda Sonia ArliestaBelum ada peringkat
- OPTIMALKAN GINJAL DAN CAIRAN TUBUHDokumen5 halamanOPTIMALKAN GINJAL DAN CAIRAN TUBUHSigit NurawalinBelum ada peringkat
- LTM Pemicu 1 Modul Ginjal Dan Cairan TubuhDokumen7 halamanLTM Pemicu 1 Modul Ginjal Dan Cairan TubuhWahyu PermatasariBelum ada peringkat
- LO Body Liquid RegulationsDokumen25 halamanLO Body Liquid Regulationsmol3yBelum ada peringkat
- Pengenceran dan Pemekatan UrinDokumen2 halamanPengenceran dan Pemekatan UrinSuhayatra Putra75% (4)
- Pengaturan Osmolaritas Cairan Ekstrasel Dan Konsentrasi NatriumDokumen2 halamanPengaturan Osmolaritas Cairan Ekstrasel Dan Konsentrasi Natriumn u r m a100% (2)
- Fisiologi Anti Diuretic HormoneDokumen22 halamanFisiologi Anti Diuretic HormonewedanurdayatBelum ada peringkat
- Pengaturan Osmolaritas Cairan Ekstra Seluler Dan Konsentrasi NatriumDokumen14 halamanPengaturan Osmolaritas Cairan Ekstra Seluler Dan Konsentrasi NatriumDago DagoBelum ada peringkat
- IKD 4, Kebutuhan Cairan ElektrolitDokumen13 halamanIKD 4, Kebutuhan Cairan ElektrolitRanaSoranaBelum ada peringkat
- Laporan Praktikum Fisiologi Kelompok A1Dokumen31 halamanLaporan Praktikum Fisiologi Kelompok A1Ivo AfianiBelum ada peringkat
- KONSENTRASI URINDokumen22 halamanKONSENTRASI URINAnnisBelum ada peringkat
- Keseimbangan Cairan, Elektrolit, Dan Asam BasaDokumen21 halamanKeseimbangan Cairan, Elektrolit, Dan Asam Basarini octavia33% (3)
- Kelompok 5 Kelas B 2016Dokumen25 halamanKelompok 5 Kelas B 2016kiki ikrimaBelum ada peringkat
- MEKANISME COUNTERCURRENT MULTIPLIERDokumen2 halamanMEKANISME COUNTERCURRENT MULTIPLIERAndreas JonathanBelum ada peringkat
- Tugas NEURODokumen14 halamanTugas NEUROEtwien ResKinta PauLusBelum ada peringkat
- Mekanisme Pengaturan Kadar Air TubuhDokumen2 halamanMekanisme Pengaturan Kadar Air TubuhChrisBelum ada peringkat
- Klara Kapitan Tugas Farmakologi VetDokumen2 halamanKlara Kapitan Tugas Farmakologi VetKlara KapitanBelum ada peringkat
- REFLEKS BARORESEPTOR DAN LFGDokumen3 halamanREFLEKS BARORESEPTOR DAN LFGFantastik4 4Belum ada peringkat
- Pengaturan Faal Cairan Dan ElektrolitDokumen2 halamanPengaturan Faal Cairan Dan Elektrolitlily putih0% (2)
- Keseimbangan Cairan dan ElektrolitDokumen5 halamanKeseimbangan Cairan dan ElektrolitHanny FadhilaBelum ada peringkat
- Modul Praktkum BSN 3Dokumen10 halamanModul Praktkum BSN 3Anggi Putri Ariyani100% (1)
- 12 EdemaDokumen15 halaman12 EdemaHumala Prika Aditama100% (1)
- Pengenceran Dan Pemekatan UrinDokumen2 halamanPengenceran Dan Pemekatan UrinFarah MuthiaBelum ada peringkat
- ELEKTROLITDokumen9 halamanELEKTROLITReza SaraswatiBelum ada peringkat
- Keseimbangan Cairan Dan Osmolaritas Cairan EkstraselulerDokumen51 halamanKeseimbangan Cairan Dan Osmolaritas Cairan EkstraseluleriikBelum ada peringkat
- Proses Reasorbsi Dan Sekresi Di Duktus KoligentesDokumen16 halamanProses Reasorbsi Dan Sekresi Di Duktus KoligentesRakhmad TriharsadiBelum ada peringkat
- Kehausan HipovolemikDokumen6 halamanKehausan HipovolemikAtika LailanaBelum ada peringkat
- Diuresis Modul Ginjal dan Cairan TubuhDokumen31 halamanDiuresis Modul Ginjal dan Cairan Tubuhdiah poppy utamiBelum ada peringkat
- Osmoregulasi 2Dokumen6 halamanOsmoregulasi 2LaluhermawanBelum ada peringkat
- CHATERINE - ANESTESIDokumen17 halamanCHATERINE - ANESTESISamuel Shephard SupitBelum ada peringkat
- Kelompok 5 - Kelas B - 2016Dokumen29 halamanKelompok 5 - Kelas B - 2016kiki ikrimaBelum ada peringkat
- Sistem RAADokumen8 halamanSistem RAAMila Nur AiniBelum ada peringkat
- Keseimbangan Cairan Tubuh - UchiDokumen4 halamanKeseimbangan Cairan Tubuh - UchiAnisa Trianti100% (1)
- Anatomi Dan FisiologiDokumen26 halamanAnatomi Dan FisiologiRosela Hibiscus SabdariffaBelum ada peringkat
- Latar Belakang Dan Tujuan Praktikum RenalDokumen6 halamanLatar Belakang Dan Tujuan Praktikum RenalANGGA PRABAWABelum ada peringkat
- OPTIMASI GINJALDokumen8 halamanOPTIMASI GINJALPuji RahayuBelum ada peringkat
- LP CKD + HDDokumen49 halamanLP CKD + HDTri Medyan PrasetyoBelum ada peringkat
- Etiologi TrichinelliasisDokumen2 halamanEtiologi TrichinelliasisGrecia SunurBelum ada peringkat
- AsdfghjDokumen8 halamanAsdfghjTirza Stephanie Nggaluama PandieBelum ada peringkat
- Fisiologi MiksiDokumen3 halamanFisiologi MiksiTirza Stephanie Nggaluama PandieBelum ada peringkat
- Klasifikasi Syok HipovolemikDokumen2 halamanKlasifikasi Syok HipovolemikTirza Stephanie Nggaluama PandieBelum ada peringkat
- Masalah Yang Sering Ditimbulkan Akibat Insomnia Pada LansiaDokumen1 halamanMasalah Yang Sering Ditimbulkan Akibat Insomnia Pada LansiaTirza Stephanie Nggaluama PandieBelum ada peringkat
- Sitii Khadija InkontinesDokumen1 halamanSitii Khadija InkontinesTirza Stephanie Nggaluama PandieBelum ada peringkat
- Amo Xixili NDokumen2 halamanAmo Xixili NTirza Stephanie Nggaluama PandieBelum ada peringkat
- Anatomi Sendi PanggulDokumen6 halamanAnatomi Sendi PanggulTirza Stephanie Nggaluama Pandie100% (1)
- Definisi Dan Jenis LukaDokumen4 halamanDefinisi Dan Jenis LukaAnastasia BhatoBelum ada peringkat
- CSL AntropometriDokumen6 halamanCSL AntropometriGrecia SunurBelum ada peringkat
- IMUNISASI-OSCEDokumen3 halamanIMUNISASI-OSCEGrecia SunurBelum ada peringkat
- GeriatriDokumen9 halamanGeriatriTirza Stephanie Nggaluama PandieBelum ada peringkat
- Tahapan Makan IchaDokumen7 halamanTahapan Makan IchaTirza Stephanie Nggaluama PandieBelum ada peringkat
- Hubungan Obat Dengan Riwayat Penyakit PoppyDokumen1 halamanHubungan Obat Dengan Riwayat Penyakit PoppyTirza Stephanie Nggaluama PandieBelum ada peringkat
- Efek Alkohol Terhadap TubuhDokumen4 halamanEfek Alkohol Terhadap TubuhTirza Stephanie Nggaluama PandieBelum ada peringkat
- Skor Apgar BabyDokumen29 halamanSkor Apgar BabyYanaBelum ada peringkat
- GeriatriDokumen9 halamanGeriatriTirza Stephanie Nggaluama PandieBelum ada peringkat
- CiriDokumen3 halamanCiriTirza Stephanie Nggaluama PandieBelum ada peringkat
- BRUSELOSISDokumen6 halamanBRUSELOSISTirza Stephanie Nggaluama PandieBelum ada peringkat
- GeriatriDokumen9 halamanGeriatriTirza Stephanie Nggaluama PandieBelum ada peringkat
- Cacing Tambang (Hookworm) PraktikumDokumen17 halamanCacing Tambang (Hookworm) PraktikumTirza Stephanie Nggaluama PandieBelum ada peringkat
- Tugas Tik PetrikDokumen7 halamanTugas Tik PetrikTirza Stephanie Nggaluama PandieBelum ada peringkat
- BRUSELOSISDokumen6 halamanBRUSELOSISTirza Stephanie Nggaluama PandieBelum ada peringkat
- Kelompok 3 FarmakologiDokumen20 halamanKelompok 3 FarmakologiTirza Stephanie Nggaluama PandieBelum ada peringkat
- UPT PerpustakaanDokumen4 halamanUPT PerpustakaanTirza Stephanie Nggaluama PandieBelum ada peringkat
- MINOSIKLIN DAN TETRASIKLINDokumen4 halamanMINOSIKLIN DAN TETRASIKLINTirza Stephanie Nggaluama PandieBelum ada peringkat
- Tugas Tik PetrikDokumen7 halamanTugas Tik PetrikTirza Stephanie Nggaluama PandieBelum ada peringkat
- InternetDokumen7 halamanInternetTirza Stephanie Nggaluama PandieBelum ada peringkat
- Lepto Spiros IsDokumen11 halamanLepto Spiros IsTirza Stephanie Nggaluama PandieBelum ada peringkat
- Makalah Penyakit Kronik TuberculosisDokumen23 halamanMakalah Penyakit Kronik TuberculosisRahMat Yondaime TampanBelum ada peringkat