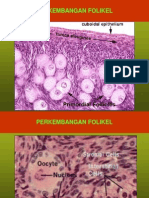
Anda mungkin juga menyukai
- Peristiwa FertilisasiDokumen6 halamanPeristiwa FertilisasiFeliciaOctofinnaBelum ada peringkat
- Embrio FertilisasiiiiiiiDokumen15 halamanEmbrio FertilisasiiiiiiiWahyuni ParamitaBelum ada peringkat
- Makalah FertilisasiDokumen37 halamanMakalah FertilisasiAinulpratamaBelum ada peringkat
- Makalah Perkembangan HewanDokumen13 halamanMakalah Perkembangan HewanReskkajuliBelum ada peringkat
- Makalah Embriologi Kelompok 2 Tentang FertilisasiDokumen19 halamanMakalah Embriologi Kelompok 2 Tentang FertilisasiIndah jefysaBelum ada peringkat
- Makalah Fertilisasi FixDokumen9 halamanMakalah Fertilisasi FixJesika RameBelum ada peringkat
- PembuahanDokumen4 halamanPembuahanNurul HabibahBelum ada peringkat
- 3 - FertilisasiDokumen13 halaman3 - FertilisasiZuhraBelum ada peringkat
- 2 Fertilisasi Pada MamaliaDokumen20 halaman2 Fertilisasi Pada MamaliaSiti ElianaBelum ada peringkat
- Jazuli Azra MR - 2009511022 - ADokumen3 halamanJazuli Azra MR - 2009511022 - A2009511022 Jazuli Azra MrBelum ada peringkat
- KLP 5 - FertilisasiDokumen14 halamanKLP 5 - FertilisasiIga Ayu MawarniBelum ada peringkat
- Kel.4 FertilisasiDokumen12 halamanKel.4 FertilisasiJesika RameBelum ada peringkat
- Makalah FertilisasiDokumen11 halamanMakalah FertilisasiLelly Luckitasari100% (2)
- Laporan Praktikum Reproduksi Dan Perkembangan Hewan - Fertilisasi MencitDokumen18 halamanLaporan Praktikum Reproduksi Dan Perkembangan Hewan - Fertilisasi MencitRifki Muhammad Iqbal100% (2)
- FERTILISASI PPT - En.idDokumen20 halamanFERTILISASI PPT - En.idHegert SitumorangBelum ada peringkat
- Fertilisasi - Embriologi - Tumbuh Kembang Janin - Dosen Lenny I SDokumen29 halamanFertilisasi - Embriologi - Tumbuh Kembang Janin - Dosen Lenny I Stiffani lorenzaBelum ada peringkat
- Laporan Pratikum Kelompok 1Dokumen5 halamanLaporan Pratikum Kelompok 1Faimat SholihinBelum ada peringkat
- BAB I FertilisasiDokumen15 halamanBAB I FertilisasiTutut TutBelum ada peringkat
- FertilisasiDokumen42 halamanFertilisasiMuhammad Aditya PermanaBelum ada peringkat
- Resume Fertilisasi - Moh. Adam Abd - 1182060068Dokumen4 halamanResume Fertilisasi - Moh. Adam Abd - 1182060068Liya Nur FatimahBelum ada peringkat
- Resume Fertilitas Kelompok 1Dokumen6 halamanResume Fertilitas Kelompok 1Fajri AkbarBelum ada peringkat
- Kelompok 2-FertilisasiDokumen19 halamanKelompok 2-FertilisasiPutri EkasBelum ada peringkat
- FertilisasiDokumen18 halamanFertilisasiSiti Nur HalizaBelum ada peringkat
- Modul 7Dokumen17 halamanModul 7Edhgar KoyBelum ada peringkat
- FertilisasiDokumen12 halamanFertilisasiMuhammad S. DizaBelum ada peringkat
- SPH II Fertilisasi-1Dokumen10 halamanSPH II Fertilisasi-1nirman-ganiBelum ada peringkat
- Fertilisasi, Fusi, SingamiDokumen15 halamanFertilisasi, Fusi, SingamiPrahesty Nur HandayaniBelum ada peringkat
- Makalah FertilisasiDokumen14 halamanMakalah FertilisasiFrangklin BarapaBelum ada peringkat
- Makalah FertilisasiDokumen14 halamanMakalah FertilisasiFrangklin BarapaBelum ada peringkat
- Fertilisasi BabiDokumen9 halamanFertilisasi BabiSendi MaulanaBelum ada peringkat
- Tr2 - PSB 20 D - Rosmida ValentinaDokumen4 halamanTr2 - PSB 20 D - Rosmida Valentinarosmidavalentina 1505Belum ada peringkat
- Makalah Embriologi Tentang Fertilisasi, Implantasi, Dan Perkembangan JaninDokumen15 halamanMakalah Embriologi Tentang Fertilisasi, Implantasi, Dan Perkembangan Janinetha rananmase50% (2)
- Laporan Penetrasi, Fertilisasi, Dan Perkembangan EmbrioDokumen8 halamanLaporan Penetrasi, Fertilisasi, Dan Perkembangan EmbrioMarisa FitrianaBelum ada peringkat
- PAPER PembuahanDokumen5 halamanPAPER PembuahanSitti salsabila MonoarfaBelum ada peringkat
- LAPORAN PenetrasiDokumen11 halamanLAPORAN PenetrasiFaishal BluesBelum ada peringkat
- Proses Fertilisasi Pada SapiDokumen10 halamanProses Fertilisasi Pada SapiNur Alfianita N100% (1)
- Fertilisasi KLP 2Dokumen15 halamanFertilisasi KLP 2Juwaeria Nur AprilyantiBelum ada peringkat
- Makalah Perkembangan Embrio AmphibiDokumen19 halamanMakalah Perkembangan Embrio AmphibiTasafima TesariBelum ada peringkat
- Soal FertilisasiDokumen4 halamanSoal FertilisasiEndahSekarBelum ada peringkat
- Makalah Embriologi Tentang Fertilisasi Implantasi Dan Perkembangan JaninDokumen15 halamanMakalah Embriologi Tentang Fertilisasi Implantasi Dan Perkembangan JaninArtheswara SidhajatiBelum ada peringkat
- FERTILISASIDokumen6 halamanFERTILISASISitiRobiahAl-adawiyahBelum ada peringkat
- FertilisasiDokumen41 halamanFertilisasinaimatul wardahBelum ada peringkat
- Fertilisasi HasnidarDokumen8 halamanFertilisasi HasnidarAksa KasimBelum ada peringkat
- Penyimpangan Pembuahan - SPHDokumen9 halamanPenyimpangan Pembuahan - SPHApsari ArgiantiBelum ada peringkat
- Bbc1-k10 Ovulasi Dan Fertilisasi 16-9Dokumen31 halamanBbc1-k10 Ovulasi Dan Fertilisasi 16-9Anika Restu PradiniBelum ada peringkat
- PBL Blok 4Dokumen14 halamanPBL Blok 4jimbon amnifuBelum ada peringkat
- Modul Fertilisasi AwalDokumen31 halamanModul Fertilisasi AwalLidya YanuartaBelum ada peringkat
- Bab I Fertilisasi VertebrataDokumen15 halamanBab I Fertilisasi VertebrataAnisa Rizky AmaliaBelum ada peringkat
- Makalah Perkembangan Hewan FertilisasiDokumen14 halamanMakalah Perkembangan Hewan FertilisasiAprilia Ramadayani100% (2)
- Tugas 1Dokumen6 halamanTugas 1wahyuBelum ada peringkat
- KesproDokumen14 halamanKesproLyena El-chamdBelum ada peringkat
- Tugas PERAWAN FERTILISASI DAN GASTRULASIDokumen17 halamanTugas PERAWAN FERTILISASI DAN GASTRULASINita Dapa OleBelum ada peringkat
- Soal Perk. Hewan PutriDokumen2 halamanSoal Perk. Hewan PutriPutri AliyahBelum ada peringkat
- FertilisasiDokumen12 halamanFertilisasiAzhar Edy WiranataBelum ada peringkat
- Fertilisasi 2010Dokumen56 halamanFertilisasi 2010Didin Puspita100% (1)
- Kelompok 9 - LKM FertilisasiDokumen6 halamanKelompok 9 - LKM FertilisasiNahidah AdillafarBelum ada peringkat
- Makalah PBL Embriologi Blok 4Dokumen10 halamanMakalah PBL Embriologi Blok 4MarthaLhtBelum ada peringkat
- Tugas SPH Sistem ReproduksiDokumen3 halamanTugas SPH Sistem ReproduksiHendrawan OyzBelum ada peringkat
- Mikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaDari EverandMikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaPenilaian: 2.5 dari 5 bintang2.5/5 (2)
- Materi SBDP Poster Dan LKDokumen6 halamanMateri SBDP Poster Dan LKHanifBelum ada peringkat
- LK IPA 4.5 MagnetDokumen1 halamanLK IPA 4.5 MagnetHanifBelum ada peringkat
- RPP Sistem PencernaanDokumen21 halamanRPP Sistem PencernaanHanif100% (2)
- Keajaiban Lampu PijarDokumen14 halamanKeajaiban Lampu PijarHanifBelum ada peringkat
- HermaproditDokumen13 halamanHermaproditHanifBelum ada peringkat
- Prak 1 Sedotan Ajaib & ElektroskopDokumen18 halamanPrak 1 Sedotan Ajaib & ElektroskopHanifBelum ada peringkat